Biogeochemical Responses to Holocene Catchment-Lake
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Glacial Map of Nw
TASMANI A DEPARTMENT OF MIN ES GEOLOGICAL SURV EY RECORD No.6 .. GLACIAL MAP OF N.W. - CEN TRAL TASMANIA by Edward Derbyshire Issued under the authority of The Honourable ERIC ELLIOTT REECE, M.H.A. , Minister for Mines for Tasmania ......... ,. •1968 REGISTERED WITH G . p.a. FOR TRANSMISSION BY POST A5 A 800K D. E . WIL.KIN SOS. Government Printer, Tasmania 2884. Pr~ '0.60 PREFACE In the published One Mile Geological Maps of the Mackintosh. Middlesex, Du Cane and 8t Clair Quadrangles the effects of Pleistocene glaciation have of necessity been only partially depicted in order that the solid geology may be more clearly indicated. However, through the work of many the region covered by these maps and the unpublished King Wi11 iam and Murchison Quadrangles is classic both throughout AustraHa and Overseas because of its modification by glaciation. It is, therefore. fitting that this report of the most recent work done in the region by geomorphology specialist, Mr. E. Derbyshire, be presented. J. G. SYMONS, Director of Mmes. 1- CONTENTS PAGE INTRODUCTION 11 GENERAL STR UCT UIIE AND MOIIPHOLOGY 12 GLACIAL MORPHOLOGY 13 Glacial Erosion ~3 Cirques 14 Nivation of Cirques 15 Discrete Glacial Cirques 15 Glacial Valley-head Cirques 16 Over-ridden Cirques 16 Rock Basin s and Glacial Trou~hs 17 Small Scale Erosional Effects 18 Glacial Depositional Landforms 18 GLACIAL SEDIMENTS 20 Glacial Till 20 Glacifluvial Deposits 30 Glacilacustrine Deposits 32 STIIATIGIIAPHY 35 REFERENCES 40 LIST OF FIGURES PAGE Fig. 1. Histogram showing orientation of the 265 cirques shown on the Glacial Map 14 Fig. -

A Survey of the Nation's Lakes
A Survey of the Nation’s Lakes – EPA’s National Lake Assessment and Survey of Vermont Lakes Vermont Agency of Natural Resources Department of Environmental Conservation - Water Quality Division 103 South Main 10N Waterbury VT 05671-0408 www.vtwaterquality.org Prepared by Julia Larouche, Environmental Technician II January 2009 Table of Contents List of Tables and Figures........................................................................................................................................................................... ii Introduction................................................................................................................................................................................................. 1 What We Measured..................................................................................................................................................................................... 4 Water Quality and Trophic Status Indicators.......................................................................................................................................... 4 Acidification Indicator............................................................................................................................................................................ 4 Ecological Integrity Indicators................................................................................................................................................................ 4 Nearshore Habitat Indicators -

2007 Annual Report
Hydro Tasmania Annual Report 07 Australia’s leading renewable energy business Achievements & Challenges for 2006/07 Achievements Ensuring Utilising Basslink Profit after tax Returns to Sale of Bell Bay Capital Further investment Targeted cost Slight increase in Hydro Tasmania Hydro Tasmania Integration of continuity of helps manage low of $79.4 million; Government of power site and gas expenditure of in Roaring 40s of reduction program staff engagement Consulting office Consulting sustainability electricity supply water storages underlying $57.8 million turbines to Alinta $54.2 million, $10 million as joint realises recurrent with Hydro opened in New achieved national performance to Tasmania in profit of • Dividend including Gordon venture builds savings of Tasmania among Delhi success as part reporting time of drought $19.5 million $21.2 million Power Station wind portfolio in $7.7 million the better of bid to receive better reflects • Income tax redevelopment Australia, China performing an $8.7 million operating result equivalent and Tungatinah and India businesses grant for a major and takes account $28.7 million switchyard nationally water monitoring of impact of low • Loan guarantee upgrade project inflows fee $5.1 million • Rates equivalent $2.8 million Challenges Operational and financial Protection of water Environmental risks Restructuring the Business response to Improving safety Increased greenhouse The direction of national Continuous improvement pressures as a result of storages as levels dipped as a result of low business -

Light-Induced Changes in Fatty Acid Profiles of Specific Lipid Classes in Several Freshwater Phytoplankton Species
Light-induced changes in fatty acid profiles of specific lipid classes in several freshwater phytoplankton species Alexander Wacker1*, Maike Piepho2, John L. Harwood3, Irina A. Guschina3, Michael T. Arts4 1Institute of Biology and Biochemistry, University of Potsdam, Germany, 2Institute of 3 Aquatic Ecology, University of Rostock, Germany, School of Biosciences, Cardiff University, United Kingdom, 4Department of Chemistry and Biology, Ryerson University, Canada Submitted to Journal: Frontiers in Plant Science Specialty Section: Plant Metabolism and Chemodiversity ISSN: 1664-462X Article type: Original Research Article Received on: 23 Dec 2015 Accepted on: 19 Feb 2016 Provisional PDF published on: Provisional19 Feb 2016 Frontiers website link: www.frontiersin.org Citation: Wacker A, Piepho M, Harwood JL, Guschina IA and Arts MT(2016) Light-induced changes in fatty acid profiles of specific lipid classes in several freshwater phytoplankton species. Front. Plant Sci. 7:264. doi:10.3389/fpls.2016.00264 Copyright statement: © 2016 Wacker, Piepho, Harwood, Guschina and Arts. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution and reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms. This Provisional PDF corresponds to the article as it appeared upon acceptance, after peer-review. Fully formatted PDF and full text (HTML) versions will be made available soon. Frontiers in Plant Science | www.frontiersin.org Provisional 1 Light-induced changes in fatty acid profiles of specific lipid classes in several 2 freshwater phytoplankton species 3 4 5 Alexander Wacker1*, Maike Piepho2, John L. -

World Heritage Values and to Identify New Values
FLORISTIC VALUES OF THE TASMANIAN WILDERNESS WORLD HERITAGE AREA J. Balmer, J. Whinam, J. Kelman, J.B. Kirkpatrick & E. Lazarus Nature Conservation Branch Report October 2004 This report was prepared under the direction of the Department of Primary Industries, Water and Environment (World Heritage Area Vegetation Program). Commonwealth Government funds were contributed to the project through the World Heritage Area program. The views and opinions expressed in this report are those of the authors and do not necessarily reflect those of the Department of Primary Industries, Water and Environment or those of the Department of the Environment and Heritage. ISSN 1441–0680 Copyright 2003 Crown in right of State of Tasmania Apart from fair dealing for the purposes of private study, research, criticism or review, as permitted under the Copyright Act, no part may be reproduced by any means without permission from the Department of Primary Industries, Water and Environment. Published by Nature Conservation Branch Department of Primary Industries, Water and Environment GPO Box 44 Hobart Tasmania, 7001 Front Cover Photograph: Alpine bolster heath (1050 metres) at Mt Anne. Stunted Nothofagus cunninghamii is shrouded in mist with Richea pandanifolia scattered throughout and Astelia alpina in the foreground. Photograph taken by Grant Dixon Back Cover Photograph: Nothofagus gunnii leaf with fossil imprint in deposits dating from 35-40 million years ago: Photograph taken by Greg Jordan Cite as: Balmer J., Whinam J., Kelman J., Kirkpatrick J.B. & Lazarus E. (2004) A review of the floristic values of the Tasmanian Wilderness World Heritage Area. Nature Conservation Report 2004/3. Department of Primary Industries Water and Environment, Tasmania, Australia T ABLE OF C ONTENTS ACKNOWLEDGMENTS .................................................................................................................................................................................1 1. -

Reimagining the Visitor Experience of Tasmania's Wilderness World
Reimagining the Visitor Experience of Tasmania’s Wilderness World Heritage Area Ecotourism Investment Profile Reimagining the Visitor Experience of Tasmania’s Wilderness World Heritage Area: Ecotourism Investment Profile This report was commissioned by Tourism Industry Council Tasmania and the Cradle Coast Authority, in partnership with the Tasmanian Government through Tourism Tasmania and the Tasmanian Parks and Wildlife Service. This report is co-funded by the Australian Government under the Tourism Industry Regional Development Fund Grants Programme. This report has been prepared by EC3 Global, TRC Tourism and Tourism Industry Council Tasmania. Date prepared: June 2014 Design by Halibut Creative Collective. Disclaimer The information and recommendations provided in this report are made on the basis of information available at the time of preparation. While all care has been taken to check and validate material presented in this report, independent research should be undertaken before any action or decision is taken on the basis of material contained in this report. This report does not seek to provide any assurance of project viability and EC3 Global, TRC Tourism and Tourism Industry Council Tasmania accept no liability for decisions made or the information provided in this report. Cover photo: Huon Pine Walk Corinna The Tarkine - Rob Burnett & Tourism Tasmania Contents Background...............................................................2 Reimagining the Visitor Experience of the TWWHA .................................................................5 -

Appendix Page
NORTH CAROLINA DEPARTMENT OF ENVIRONMENT AND NATURAL RESOURCES Division of Water Quality Environmental Sciences Section August 2004 This page was intentionally left blank NCDENR, Division of Water Quality Basinwide Assessment Report – Cape Fear River Basin - August 2004 1 TABLE OF CONTENTS Page LIST OF APPENDICIES ........................................................................................................................ 5 LIST OF TABLES................................................................................................................................... 7 LIST OF FIGURES .............................................................................................................................. 11 OVERVIEW OF THE WATER QUALITY OF THE CAPE FEAR RIVER BASIN.....................................17 EXECUTIVE SUMMARIES BY PROGRAM AREA.................................................................................27 FISHERIES ...................................................................................................................................... 27 BENTHIC MACROINVERTEBRATES............................................................................................. 30 LAKE ASSESSMENT....................................................................................................................... 32 PHYTOPLANKTON MONITORING................................................................................................. 33 AMBIENT MONITORING................................................................................................................ -

Western Lakes Fishery Management Plan 2002
Wilderness Fishery Western Lakes Western November 2002 F ISHERY M ANAGEMENT P LAN WESTERN LAKES WESTERN LAKES F ISHERY M ANAGEMENT P LAN November 2002 November 2002 Western Lakes – Fishery Management Plan Western Lakes – Fishery Management Plan November 2002 Executive Summary Introduction This fishery management plan is a subsidiary plan under the Tasmanian Wilderness World Heritage Area Management Plan (WHA plan). The plan covers all areas of responsibility for which the Inland Fisheries Service (IFS) has statutory control; freshwater native species, freshwater recreational fisheries, and freshwater commercial fisheries. The plan also makes several recommendations on land management issues for consideration by the Parks and Wildlife Service (PWS). The area covered by the plan includes the Central Plateau Conservation Area west of the Lake Highway and the Walls of Jerusalem National Park, both of which lie within the Tasmanian Wilderness World Heritage Area. Environment This section examines the impacts of users (primarily anglers) on the environment and specifically water quality, and how these impacts can be minimised while maintaining angling opportunities. Management prescriptions focus on monitoring and review of water quality and the impacts of boating, wading and weir construction in various waters, and where necessary, implementing remediation measures. An information and education approach with the particular emphasis on the use of signage, will play an important role. Establishment of alternative boating access outside of the Western Lakes, development of a boating code of practice and review of current boating regulations will assist in minimising boating impacts. Additionally, the IFS will encourage and support studies that examine the impacts of boating and wading. -
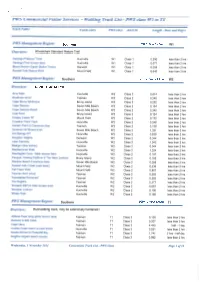
Walking Track List - PWS Class Wl to T4
PWS Commercial Visitor Services - Walking Track List - PWS class Wl to T4 Track Name FieldCentre PWS class AS2156 Length - Kms and Days PWS Management Region: Southern PWS Track Class: VV1 Overview: Wheelchair Standard Nature Trail Hastings Platypus Track Huonville W1 Class 1 0.290 less than 2 hrs Hastings Pool access track Huonville W1 Class 1 0.077 less than 2 hrs Mount Nelson Signal Station Tracks Derwent W1 Class 1 0.059 less than 2 hrs Russell Falls Nature Walk Mount Field W1 Class 1 0.649 less than 2 hrs PWS Management Region: Southern PWS Track Class: W2 Overview: Standard Nature Trail Arve Falls Huonville W2 Class 2 0.614 less than 2 hrs Blowhole circuit Tasman W2 Class 2 0.248 less than 2 hrs Cape Bruny lighthouse Bruny Island W2 Class 2 0.252 less than 2 hrs Cape Deslacs Seven Mile Beach W2 Class 2 0.154 less than 2 hrs Cape Deslacs Beach Seven Mile Beach W2 Class 2 0.345 less than 2 hrs Coal Point Bruny Island W2 Class 2 0.124 less than 2 hrs Creepy Crawly NT Mount Field W2 Class 2 0.175 less than 2 hrs Crowther Point Track Huonville W2 Class 2 0.248 less than 2 hrs Garden Point to Carnarvon Bay Tasman W2 Class 2 3.138 less than 2 hrs Gordons Hill fitness track Seven Mile Beach W2 Class 2 1.331 less than 2 hrs Hot Springs NT Huonville W2 Class 2 0.839 less than 2 hrs Kingston Heights Derwent W2 Class 2 0.344 less than 2 hrs Lake Osbome Huonville W2 Class 2 1.042 less than 2 hrs Maingon Bay lookout Tasman W2 Class 2 0.044 less than 2 hrs Needwonnee Walk Huonville W2 Class 2 1.324 less than 2 hrs Newdegate Cave - Main access -

Trophic Classification of Selected Colorado Lakes
JPL PUBLICATION 78-100 EPA-600/4-79-005 Trophic Classification of Selected Colorado Lakes Ribhard J. Black-well Jet Propulsion Laboratory' Dale H. P. Boland U.S, EnvironmentahProtection Agency (NASA-CR-158500) TROPHIC CLASSIFICATION OF N79-22591 SELECTED COLORADO LAKES (Jet Propulsion Lab.) 210 p HC A10/HF A01 CSCL 08H Unclas G3/43 25085 January 197 SRIGINAL CONTAIRS 1Ol.R ILLUSMTr)h$s 'Prepared for - National Aer-onutics and Space Administration and U.S. Environmental Protection Agency Jet Propulsion Laboratory 4c, California, Institute of Technology Pasadena, California SUBJECT: Errata Please note the following corrections to JPL Publication 78-100, Trophic Classification of Selected Colorado Lakes, by Richard J. Blackwell and Dale H. P. Boland, dated January 1979: Page 5-1, paragraph (2): Line 1, change "MSS and MSS" to "MKS and MSS" Line 5, change "MSS-related" to "MMS-related" JPL PUBLICATION 78-100 tA-b00/4h79-005 TROPHIC CLASSIFICATION OF SELECTED COLORADO LAKES Lake Classification Through the Amalgamation of Contact-Sensed Data and Digitally Processed Multispectral Scanner Data Acquired by Satellite and Aircraft Richard J. Blackwell Dale H. P. Boland Earth Resources Application Group Water and Land Quality Branch Jet Propulsion Laboratory Environmental Monitoring and California Institute of Technology Support Laboratory - Las Vegas Pasadena, California National Aeronautics and U.S. Environmental Protection Agency Space Administration Office of Research and Development Washington, D.C. 20546 Las Vegas, Nevada 89114 January 1979 The research described in this report was carried out by the Jet Propulsion Laboratory, California Institute of Technology, together with the U. S. Environmental Protection Agency and was jointly sponsored by the National Aeronautics and Space Administration and the U. -

Dove Lake Viewing Shelter Construction 20 July 2021
TASMANIA PARKS AND WILDLIFE SERVICE FAQ Dove Lake Viewing Shelter Construction 20 July 2021 Q1. What is happening at Dove Lake? The construction of the Dove Lake viewing platform is part of a $160 million plan to enhance the Cradle Mountain visitor experience with new infrastructure and amenities. This is the second stage of the overall Cradle Mountain Master Plan redevelopment. The viewing platform has been designed to blend into the Cradle Mountain landscape and features earthy colours, seating and wall-to-ceiling glass panels, with its design to fit within the existing car park footprint to minimise the impact on the natural environment and will be setback from the edge of Dove Lake. The structure will be joined by a new bus shelter and amenities building, outdoor viewing area, parking, turning circle and connections to the Dove Lake walking tracks and Glacier Rock viewing platform. Construction commenced in April 2020 and is due for completion in summer 2021. Q2. Is vehicle access to Dove Lake about to change? All vehicle access to Dove Lake will be restricted during the construction of the Dove Lake Viewing Shelter, within the Cradle Mountain-Lake St Clair National Park and Tasmanian Wilderness World Heritage Area. Visitors wanting to access Dove Lake for short day walks will not be inconvenienced as the shuttle bus service will continue to operate to a very frequent timetable during the peak periods. During construction, temporary infrastructure (bus turning area, toilets and temporary walking tracks) will be in place at Dove Lake from early 2020. Space for site offices and construction vehicles will result in no parking being available. -

Light-Induced Changes in Fatty Acid Profiles of Specific Lipid
ORIGINAL RESEARCH published: 16 March 2016 doi: 10.3389/fpls.2016.00264 Light-Induced Changes in Fatty Acid Profiles of Specific Lipid Classes in Several Freshwater Phytoplankton Species Alexander Wacker 1*, Maike Piepho 2, John L. Harwood 3, Irina A. Guschina 3 and Michael T. Arts 4 1 Theoretical Aquatic Ecology and Ecophysiology, Institute of Biochemistry and Biology, University of Potsdam, Potsdam, Germany, 2 Department for Ecology, Institute of Aquatic Ecology, University of Rostock, Rostock, Germany, 3 School of Biosciences, Cardiff University, Cardiff, UK, 4 Department of Chemistry and Biology, Ryerson University, Toronto, ON, Canada We tested the influence of two light intensities [40 and 300 µmol PAR / (m2s)] on the fatty acid composition of three distinct lipid classes in four freshwater phytoplankton species. We chose species of different taxonomic classes in order to detect potentially similar reaction characteristics that might also be present in natural phytoplankton communities. From samples of the bacillariophyte Asterionella formosa, the chrysophyte Chromulina Edited by: sp., the cryptophyte Cryptomonas ovata and the zygnematophyte Cosmarium botrytis Jeffrey Leblond, Middle Tennessee State University, we first separated glycolipids (monogalactosyldiacylglycerol, digalactosyldiacylglycerol, USA and sulfoquinovosyldiacylglycerol), phospholipids (phosphatidylcholine, Reviewed by: phosphatidylethanolamine, phosphatidylglycerol, phosphatidylinositol, and Justin Murdock, Tennessee Technological University, phosphatidylserine) as