MUC4-Promoted Neural Invasion Is Mediated by the Axon Guidance Factor Netrin-1 in PDAC
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2014 Interim Results Announcement
2014 Interim Results Presentation August 2014 CHINA EVERBRIGHT INTERNATIONAL LIMITED 中國光大國際有限公司 Forward-looking statements This presentation may contain forward-looking statements. Any such forward-looking statements are based on a number of assumptions about the operations of China Everbright International Limited (the “Company”) and factors beyond the Company's control and are subject to significant risks and uncertainties, and accordingly, actual results may differ materially from these forward-looking statements. The Company undertakes no obligation to update these forward-looking statements for events or circumstances that occur subsequent to such dates. The information in this presentation should be considered in the context of the circumstances prevailing at the time of its presentation and has not been, and will not be, updated to reflect material developments which may occur after the date of this presentation. The slides forming part of this presentation have been prepared solely as a support for oral discussion about background information about the Company. No representation or warranty, express or implied, is made as to, and no reliance should be placed on, the fairness, accuracy, completeness or correctness of any information or opinion contained herein. It should not be regarded by recipients as a substitute for the exercise of their own judgment. Information and opinion contained in this presentation may be based on or derived from the judgment and opinion of the management of the Company. Such information is not always capable of verification or validation. None of the Company or financial adviser of the Company, or any of their respective directors, officers, employees, agents or advisers shall be in any way responsible for the contents hereof, or shall be liable for any loss arising from use of the information contained in this presentation or otherwise arising in connection therewith. -

Jiangsu(PDF/288KB)
Mizuho Bank China Business Promotion Division Jiangsu Province Overview Abbreviated Name Su Provincial Capital Nanjing Administrative 13 cities and 45 counties Divisions Secretary of the Luo Zhijun; Provincial Party Li Xueyong Committee; Mayor 2 Size 102,600 km Shandong Annual Mean 16.2°C Jiangsu Temperature Anhui Shanghai Annual Precipitation 861.9 mm Zhejiang Official Government www.jiangsu.gov.cn URL Note: Personnel information as of September 2014 [Economic Scale] Unit 2012 2013 National Share (%) Ranking Gross Domestic Product (GDP) 100 Million RMB 54,058 59,162 2 10.4 Per Capita GDP RMB 68,347 74,607 4 - Value-added Industrial Output (enterprises above a designated 100 Million RMB N.A. N.A. N.A. N.A. size) Agriculture, Forestry and Fishery 100 Million RMB 5,809 6,158 3 6.3 Output Total Investment in Fixed Assets 100 Million RMB 30,854 36,373 2 8.2 Fiscal Revenue 100 Million RMB 5,861 6,568 2 5.1 Fiscal Expenditure 100 Million RMB 7,028 7,798 2 5.6 Total Retail Sales of Consumer 100 Million RMB 18,331 20,797 3 8.7 Goods Foreign Currency Revenue from Million USD 6,300 2,380 10 4.6 Inbound Tourism Export Value Million USD 328,524 328,857 2 14.9 Import Value Million USD 219,438 221,987 4 11.4 Export Surplus Million USD 109,086 106,870 3 16.3 Total Import and Export Value Million USD 547,961 550,844 2 13.2 Foreign Direct Investment No. of contracts 4,156 3,453 N.A. -

Preliminary Determination in the Antidumping Duty Investigations on Imports of Citric Acid and Certain Citrate Salts from Canada
FACT SHEET Commerce Preliminarily Finds Dumping of Citric Acid and Certain Citrate Salts from the People’s Republic of China and Canada • On November 13, the Department of Commerce (Commerce) announced its affirmative preliminary determinations in the antidumping duty (AD) investigations of imports of citric acid and certain citrate salts from the People’s Republic of China (China) and Canada. Citric acid and citrate salts are used in various food and beverage products including carbonated and non-carbonated drinks, and frozen foods, as well as laundry detergents and household cleaning products. • Dumping occurs when a foreign company sells a product in the United States at less than normal value. • Commerce preliminarily determined that Chinese and Canadian producers/exporters have sold citric acid and citrate salts in the United States at 119.41 to 156.87 percent, and 20.88 percent below normal value, respectively. • In the China investigation, mandatory respondents TTCA Co., Ltd. (a.k.a. Shandong TTCA Biochemistry Co., Ltd.) and Yixing Union Biochemical Co., Ltd., received preliminary dumping rates of 150.09 and 119.41 percent, respectively. Eleven Chinese exporters qualified for a separate rate of 134.75 percent. All other exporters will receive the China-wide rate of 156.87 percent. • In the Canada investigation, mandatory respondent Jungbunzlauer Technology GMBH & Co KG, received a preliminary dumping rate of 20.88 percent. All other Canadian exporters will receive a rate of 20.88 percent. • As a result of these preliminary determinations, Commerce will instruct U.S. Customs and Border Protection to collect a cash deposit or bond based on these preliminary rates. -

Correlation Between Peripheral White Blood Cell Counts And
Int. J. Med. Sci. 2013, Vol. 10 758 Ivyspring International Publisher International Journal of Medical Sciences 2013; 10(6):758-765. doi: 10.7150/ijms.6155 Research Paper Correlation between Peripheral White Blood Cell Counts and Hyperglycemic Emergencies Wei Xu1,2 ,∗, Hai-feng Wu3,∗, Shao-gang Ma2,∗, Feng Bai2, Wen Hu2, Yue Jin4, Hong Liu2 1. School of Medicine, Southeast University, Nanjing, Jiangsu 210009, China; 2. Department of Endocrinology, the Affiliated Huai'an Hospital of Xuzhou Medical College, Huai'an, Jiangsu 223002, China; 3. Department of Critical Care Medicine, the Affiliated Yixing People’s Hospital of Jiangsu University, Yixing, Jiangsu 214200, China; 4. Department of Clinical Laboratory, the Affiliated Huai'an Hospital of Xuzhou Medical College, Huai'an, Jiangsu 223002, China. ∗ Co-first authors. Corresponding author: Shao-gang Ma, Ph.D., M.D. Address: No. 60 South Huaihai Road, Huai'an 223002, China E-mail: [email protected] Fax/Phone: +86-517-8394 3591. © Ivyspring International Publisher. This is an open-access article distributed under the terms of the Creative Commons License (http://creativecommons.org/ licenses/by-nc-nd/3.0/). Reproduction is permitted for personal, noncommercial use, provided that the article is in whole, unmodified, and properly cited. Received: 2013.02.26; Accepted: 2013.04.10; Published: 2013.04.18 Abstract Objective: To determine the correlation between differential leukocyte counts and hypergly- cemic emergencies. Methods: Fifty patients with diabetic ketoacidosis (DKA), 50 patients with diabetic ketosis (DK), 50 non-DK diabetic patients with stable glycemic control, and 50 normal controls were enrolled. Their total and differential leukocyte counts were measured and evaluated at admission and after treatment. -

Final Determinations in the Antidumping and Countervailing
FACT SHEET Commerce Finds Unfair Dumping and Subsidization of Citric Acid and Certain Citrate Salts from the People’s Republic of China and Canada • On April 7, the Department of Commerce (Commerce) announced its affirmative final determinations in the antidumping and countervailing duty investigations on imports of citric acid and certain citrate salts from the People’s Republic of China (China) and Canada. Citric acid and citrate salts are used in various food and beverage products including carbonated and non- carbonated drinks, and frozen foods, as well as laundry detergents and household cleaning products. • Dumping occurs when a foreign company sells a product in the United States at less than normal value. Subsidies are financial assistance from foreign governments that benefit the production, manufacture, or exportation of goods. • Commerce determined that Chinese and Canadian producers/exporters have sold citric acid and citrate salts in the United States at 94.61 to 156.87 percent, and 23.21percent below normal value, respectively. • In the China investigation, mandatory respondents TTCA Co., Ltd. (a.k.a. Shandong TTCA Biochemistry Co., Ltd.) and Yixing Union Biochemical Co., Ltd., received final dumping rates of 129.08 and 94.61 percent, respectively. Eleven Chinese exporters qualified for a separate rate of 111.85 percent. All other exporters will receive the China-wide rate of 156.87 percent. • In the Canada investigation, mandatory respondent, Jungbunzlauer Canada, Inc., received a final dumping rate of 23.21 percent. All other Canadian exporters will receive a rate of 23.21 percent. • In addition, Commerce also determined that Chinese producers/exporters have received net countervailable subsidies ranging from 3.60 to 118.95 percent. -
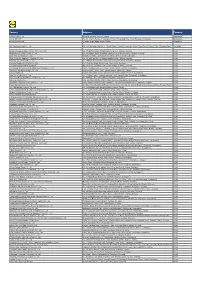
Factory Address Country
Factory Address Country Durable Plastic Ltd. Mulgaon, Kaligonj, Gazipur, Dhaka Bangladesh Lhotse (BD) Ltd. Plot No. 60&61, Sector -3, Karnaphuli Export Processing Zone, North Potenga, Chittagong Bangladesh Bengal Plastics Ltd. Yearpur, Zirabo Bazar, Savar, Dhaka Bangladesh ASF Sporting Goods Co., Ltd. Km 38.5, National Road No. 3, Thlork Village, Chonrok Commune, Korng Pisey District, Konrrg Pisey, Kampong Speu Cambodia Ningbo Zhongyuan Alljoy Fishing Tackle Co., Ltd. No. 416 Binhai Road, Hangzhou Bay New Zone, Ningbo, Zhejiang China Ningbo Energy Power Tools Co., Ltd. No. 50 Dongbei Road, Dongqiao Industrial Zone, Haishu District, Ningbo, Zhejiang China Junhe Pumps Holding Co., Ltd. Wanzhong Villiage, Jishigang Town, Haishu District, Ningbo, Zhejiang China Skybest Electric Appliance (Suzhou) Co., Ltd. No. 18 Hua Hong Street, Suzhou Industrial Park, Suzhou, Jiangsu China Zhejiang Safun Industrial Co., Ltd. No. 7 Mingyuannan Road, Economic Development Zone, Yongkang, Zhejiang China Zhejiang Dingxin Arts&Crafts Co., Ltd. No. 21 Linxian Road, Baishuiyang Town, Linhai, Zhejiang China Zhejiang Natural Outdoor Goods Inc. Xiacao Village, Pingqiao Town, Tiantai County, Taizhou, Zhejiang China Guangdong Xinbao Electrical Appliances Holdings Co., Ltd. South Zhenghe Road, Leliu Town, Shunde District, Foshan, Guangdong China Yangzhou Juli Sports Articles Co., Ltd. Fudong Village, Xiaoji Town, Jiangdu District, Yangzhou, Jiangsu China Eyarn Lighting Ltd. Yaying Gang, Shixi Village, Shishan Town, Nanhai District, Foshan, Guangdong China Lipan Gift & Lighting Co., Ltd. No. 2 Guliao Road 3, Science Industrial Zone, Tangxia Town, Dongguan, Guangdong China Zhan Jiang Kang Nian Rubber Product Co., Ltd. No. 85 Middle Shen Chuan Road, Zhanjiang, Guangdong China Ansen Electronics Co. Ning Tau Administrative District, Qiao Tau Zhen, Dongguan, Guangdong China Changshu Tongrun Auto Accessory Co., Ltd. -

Xian Road East First Road First East
ISLAND PALACE Yixing Review of Operations – Business in Mainland China Progress of Major Development Projects Changsha The Arch of Triumph (100% owned*) Changsha Songya Lake Wangxian Road East First Road East First East Fourth Road East Third Road East East Second Road East Second Tangpo Road Kaiyuan East Road Changsha National Economic-Technological Development Area The Arch of Triumph, Changsha The Arch of Triumph is a community development with around 6,700,000 square feet of premium residential units to be built in three phases. Its 33-storey Arc de Triomphe-style building is a landmark development in this new town of Xingsha. Phases 1, 2A, 2B and 3A were completed and delivered to buyers already. A new prestigious school in Changsha, namely Datong Xingsha Primary School, has also been built and become operational. The remaining phases will provide approximately 2,050,000 square feet of residential area, in addition to 320,000 square feet of serviced apartments and commercial facilities upon their successive completion during the period from 2018 to 2019. (*CIFI Holdings (Group) Co. Ltd. (“CIFI”) will participate in the development of Phases 3B and 3C and will share 30% of their costs and economic interests.) Henderson Land Development Company Limited 64 Annual Report 2017 Review of Operations – Business in Mainland China • Progress of Major Development Projects Chengdu Chengdu ICC (30% owned) Chengdu R Shuangqing o ad Chengdu Metro Line 2 Shuanggui Road D on gd a R o a d Tazishan Park 2nd Ring Road Shahepu Street Wangjianglou Guyapo Road Park Jingkang Road J Jinhua Road in g ju Yidu Ave si R o a d Lijieren Former Residence Chengdu ICC, Chengdu The 14,000,000-square-foot Chengdu ICC is a sizeable integrated development located in the business hub of Chengdu. -

Transmissibility of Hand, Foot, and Mouth Disease in 97 Counties of Jiangsu Province, China, 2015- 2020
Transmissibility of Hand, Foot, and Mouth Disease in 97 Counties of Jiangsu Province, China, 2015- 2020 Wei Zhang Xiamen University Jia Rui Xiamen University Xiaoqing Cheng Jiangsu Provincial Center for Disease Control and Prevention Bin Deng Xiamen University Hesong Zhang Xiamen University Lijing Huang Xiamen University Lexin Zhang Xiamen University Simiao Zuo Xiamen University Junru Li Xiamen University XingCheng Huang Xiamen University Yanhua Su Xiamen University Benhua Zhao Xiamen University Yan Niu Chinese Center for Disease Control and Prevention, Beijing City, People’s Republic of China Hongwei Li Xiamen University Jian-li Hu Jiangsu Provincial Center for Disease Control and Prevention Tianmu Chen ( [email protected] ) Page 1/30 Xiamen University Research Article Keywords: Hand foot mouth disease, Jiangsu Province, model, transmissibility, effective reproduction number Posted Date: July 30th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-752604/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 2/30 Abstract Background: Hand, foot, and mouth disease (HFMD) has been a serious disease burden in the Asia Pacic region represented by China, and the transmission characteristics of HFMD in regions haven’t been clear. This study calculated the transmissibility of HFMD at county levels in Jiangsu Province, China, analyzed the differences of transmissibility and explored the reasons. Methods: We built susceptible-exposed-infectious-asymptomatic-removed (SEIAR) model for seasonal characteristics of HFMD, estimated effective reproduction number (Reff) by tting the incidence of HFMD in 97 counties of Jiangsu Province from 2015 to 2020, compared incidence rate and transmissibility in different counties by non -parametric test, rapid cluster analysis and rank-sum ratio. -

Producent Adres Land
*Deze lijst bevat alle 'non-food' leveranciers die producten aan Lidl hebben geleverd in de periode tussen 1 maart 2019 en 29 februari 2020. Producent Adres Land 3W Home Fashion Heyuan Co., Ltd. Mingzhu Industrial Park, Heyuan West Road, Chuangye South Road, Heyuan, Guangdong China A.B Sales Corp. (A Unit Of Satyam Creations (Pvt) Ltd.) Plot No. 1642, Zone -09, Kolkata Leather Complex, Bantala, 24 Parganas (South), Kolkatta, West Bengal India AB Apparels Ltd. No. 225 Singair Road, Tetuljhora, Hemayetpur, Savar, Dhaka Bangladesh Above & Beyond Co., Ltd. Plot No. 116/A, 116/B, Settmu (10) Street, Myay Taing Block No. 42, Industrial Zone (1), Shwe Pyi Thar Township, Yangon Myanmar Ador Composite Ltd. 1, C&B Bazar, Gilarchala, Sreepur, Gazipur, Bd Gazipur District, Gazipur, Dhaka Bangladesh Advanced Composite Textile Ltd. Kashor Masterbari, Bhaluka, Mymensingh, Sylhet Bangladesh Afroze Textile Industries (Pvt) Ltd. Plot C-8, Scheme 33, S.I.T.E., Super Highway, Karachi, Sindh Pakistan Afroze Textile Industries (Pvt) Ltd. LA-1/A, Block 22, F.B. Area, Karachi, Sindh Pakistan Ahmed Fashions 34/1, Darus Salam Road, Dhaka Bangladesh Ai Qi Fujian Shoes Plastic Co., Ltd. Meiling Street, Shuanggou Industrial District, Sichuan, Jinjiang, Fujian China Al Hadi Textile (Pvt) Ltd. D-12 Site Super Highway Industrial Area, Karachi, Sindh Pakistan Alpine Clothings Polpithigama (Pvt) Ltd. Andarayaya, Polpithigama Sri Lanka Alpine Clothings Yapahuwa (Pvt) Ltd. Anuradhapura Road, Uduweriya, North Western Sri Lanka AMG Factory Ltd. Plot No. 51 & 52, Myay Taing Block No. 25, Shwe Lin Ban Industrial Zone, Hlaing Thar Yar Township, Yangon Myanmar Andy Accessory Co., Ltd. -

Federal Register/Vol. 80, No. 5/Thursday, January 8, 2015/Notices
1018 Federal Register / Vol. 80, No. 5 / Thursday, January 8, 2015 / Notices (Rugao Jiangbei); (18) RZBC Group Fuel. The administrative review will International Trade Commission (ITC), Shareholding Co., Ltd. (RZBC Group) continue with respect to Laiwu Taihe. the Department is issuing a and RZBC; (19) Shandong Hongshide countervailing duty (CVD) order on Assessment Chemical Co., Ltd. (Shandong carbon and certain alloy steel wire rod Hongshide); (20) Shandong TTCA The Department will instruct U.S. (steel wire rod) from the People’s Biochemistry Co., Ltd. (Shandong Customs and Border Protection (CBP) to Republic of China (PRC). Also, as TTCA); (21) Shihezi City Changyum assess countervailing duties on all explained in this notice, the Department Biochemical Co., Ltd. (Shihezi City appropriate entries. For the companies is amending its final determination to Changyum); (22) Weifang Ensign for which this review is rescinded, correct an error with respect to the Industry Co., Ltd. (Weifang Ensign); (23) countervailing duties shall be assessed identification of an affiliate of one of the Wuhan Shuangfeng Citric Acid Co., Ltd. at rates equal to the cash deposit of respondents. (Wuhan Shuangfeng); (24) Yixing Union estimated countervailing duties required DATES: Effective Date: January 8, 2015. Biochemical Co., Ltd. (Yixing Union); at the time of entry, or withdrawal from FOR FURTHER INFORMATION CONTACT: (25) Yixing Zhenfen Medical Chemical warehouse, for consumption, during the Rebecca Trainor or Reza Karamloo, Co., Ltd. (Yixing Zhenfen); and (26) period January 1, 2013, through Office II, AD/CVD Operations, Yunnan No. 2 Fuel Factory (Yunnan December 31, 2013, in accordance with Enforcement and Compliance, Fuel). 19 CFR 351.212(c)(1)(i). -

Newsletter for July 2006, Vol. 51
CHINESE CLAYART NEWSLETTER July 2006, Vol. 51 FEATURES Hands on Teapot Workshop at Wuxi Institute of Arts and Technology PROFILE Introduction to the Wuxi Institute of Arts and Technology CONFERENCE AND EVENT Q&A about the Jingdezhen Ceramic Fair and Yixing Ceramic Festival in China, October 2006 TRAVEL Tour of "Ceramic China", October tour itinerary update Letter from Ellie Bartholomew, Southern California ACTIVITIES IAC Assembly at Riga, Latvia, August 14-18, 2006 LETTERS Childhood under the Red Flag CLAY TOOL and BOOK STORE Four Kinds of Wheels for Clay Work CHINESE CLAYART " Chinese Clayart" is a newsletter emailed monthly to professional ceramic artists who want to know about ceramic art in China and things related. This newsletter will be a bridge between China and Western countries for the ceramic arts. Comments and suggestions are very welcome. (Copyright 2000, The Chinese Ceramic Art Council, USA. All rights reserved) The Chinese Ceramic Art Council, USA. P.O.Box 64392, Sunnyvale, CA 94088, USA Tel. 408-777-8319, Fax. 408-777-8321, Email: [email protected] www.chineseclayart.com Chief Editor, Guangzhen "Po" Zhou English Editor, Deborah Bouchette TOP PROFILE Introduction to the Wuxi Institute of Arts and Technology The Wuxi Institute of Arts and Technology (WIAT) is located in Yixing, the Capital of Pottery. Situated on the west bank of Taihu Lake in the delta of the Yangtze River, the city traditionally has been called "fish and rice village." It is also known as a "hometown of professors" (many Yixing- born professors teach all over the world today). The Institute covers an area of over 100 acres and its new campus building area amounts to approximately 150,000 square meters, which offers a perfect modern learning environment for more than 6,000 students. -

Disturbed Effective Connectivity Patterns in an Intrinsic Triple Network Model Are Associated with Posttraumatic Stress Disorder
Neurological Sciences (2019) 40:339–349 https://doi.org/10.1007/s10072-018-3638-1 ORIGINAL ARTICLE Disturbed effective connectivity patterns in an intrinsic triple network model are associated with posttraumatic stress disorder Yifei Weng1 & Rongfeng Qi1 & Li Zhang2 & Yifeng Luo3 & Jun Ke1 & Qiang Xu1 & Yuan Zhong1 & Jianjun Li4 & Feng Chen4 & Zhihong Cao3 & Guangming Lu 1 Received: 19 June 2018 /Accepted: 7 November 2018 /Published online: 17 November 2018 # Springer-Verlag Italia S.r.l., part of Springer Nature 2018 Abstract Background Disturbance of the triple network model was recently proposed to be associated with the occurrence of posttrau- matic stress disorder (PTSD) symptoms. Based on resting-state dynamic causal modeling (rs-DCM) analysis, we investigated the neurobiological model at a neuronal level along with potential neuroimaging biomarkers for identifying individuals with PTSD. Methods We recruited survivors of a devastating typhoon including 27 PTSD patients, 33 trauma-exposed controls (TECs), and 30 healthy controls without trauma exposure. All subjects underwent resting-state functional magnetic resonance imaging. Independent components analysis was used to identify triple networks. Detailed effective connectivity patterns were estimated by rs-DCM analysis. Spearman correlation analysis was performed on aberrant DCM parameters with clinical assessment results relevant to PTSD diagnosis. We also carried out step-wise binary logistic regression and receiver operating characteristic curve (ROC) analysis to confirm the capacity of altered effective connectivity parameters to distinguish PTSD patients. Results Within the executive control network, enhanced positive connectivity from the left posterior parietal cortex to the left dorsolateral prefrontal cortex was correlated with intrusion symptoms and showed good performance (area under the receiver operating characteristic curve = 0.879) in detecting PTSD patients.