Early Occurrence of a Pennsylvanian-Age Medullosalean Frond Similar to Alethopteris Pseudograndinioides in the Intra-Montane Basin of Bohemia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

International Organisation of Palaeobotany IOP NEWSLETTER
INTERNATIONAL UNION OF BIOLOGIC A L S C IENC ES S ECTION FOR P A L A EOBOTANY International Organisation of Palaeobotany IOP NEWSLETTER 110 August 2016 CONTENTS FROM THE SECRETARY/TREASURER IPC XIV/IOPC X 2016 IOPC 2020 IOP MEMBERSHIP IOP EXECUTIVE COMMITTEE ELECTIONS IOP WEBMASTER POSITION WHAT HAPPENED TO THE OUPH COLLECTIONS? THE PALAEOBOTANY OF ITALY UPCOMING MEETINGS CALL FOR NEWS and NOTES The views expressed in the newsletter are those of its correspondents, and do not necessarily reflect the policy of IOP. Please send us your contributions for the next edition of our newsletter (June 2016) by M ay 30th, 2016. President: Johanna Eder-Kovar (G ermany) Vice Presidents: Bob Spicer (Great Britain), Harufumi Nishida (Japan), M ihai Popa (Romania) M embers at Large: Jun W ang (China), Hans Kerp (Germany), Alexej Herman (Russia) Secretary/Treasurer/Newsletter editor: M ike Dunn (USA) Conference/Congress Chair: Francisco de Assis Ribeiro dos Santos IOP Logo: The evolution of plant architecture (© by A. R. Hemsley) I OP 110 2 August 2016 FROM THE In addition, please send any issues that you think need to be addressed at the Business SECRETARY/TREASURER meeting. I will add those to the Agenda. Dear IOP Members, Respectfully, Mike I am happy to report, that IOP seems to be on track and ready for a new Executive Council to take over. The elections are IPC XIV/IOPC X 2016 progressing nicely and I will report the results in the September/October Newsletter. The one area that is still problematic is the webmaster position. We really to talk amongst ourselves, and find someone who is willing and able to do the job. -

A New Microsporangiate Organ from the Lower Carboniferous of the Novgorod Region, Russia O
ISSN 0031-0301, Paleontological Journal, 2009, Vol. 43, No. 10, pp. 1316–1329. © Pleiades Publishing, Ltd., 2009. A New Microsporangiate Organ from the Lower Carboniferous of the Novgorod Region, Russia O. A. Orlovaa, N. R. Meyer-Melikian†,a, and N. E. Zavialovab a Moscow State University (MGU), Leninskie gory 1, Moscow, 119991 Russia b Borissiak Paleontological Institute of the Russian Academy of Sciences, ul. Profsoyuznaya 123, Moscow, 117997 Russia e-mail: [email protected] Received February 5, 2008 Abstract—A new species of the genus Telangiopsis, T. nonnae O. Orlova et Zavialova, was described on the basis of a microsporangiate organ from the Lower Carboniferous deposits of the Novgorod Region. The mor- phology of branching fertile axes, synangia, and sporangia was thoroughly studied. The three-dimensional sys- tem of fertile axes branches monopodially; ultimate axes bear numerous connivent bunches of synangia, which consist of three to six basally fused elongated ovate sporangia. The morphology and ultrastructure of prepollen grains were studied, which were extracted from the rock matrix surrounding the sporangia. The two-layered exine includes a well-developed endexine and an alveolate ectexine, with one–three rows of large thin-walled alveolae. The new species was compared with other Early Carboniferous microsporangiate organs. Key words: Early Carboniferous, Novgorod Region, fertile axis, synangia, Lyginopteridales, trilete prepollen, exine ultrastructure. DOI: 10.1134/S003103010910013X INTRODUCTION synangia and numerous casts and imprints of detached synangia were found in yellow ferruginous sandstone. During the three last decades, the interest of bota- In addition to the fertile axes, sterile remains of Lygi- nists dealing with fossil and modern plants to Carbon- nopteridales, Medullosales, and Calamopytiales were iferous synangiate pollen organs has considerably found (Orlova and Snigirevskii, 2003, 2004). -

A Stable Isotopic Investigation of Resource Partitioning Among Neosauropod Dinosaurs of the Upper Jurassic Morrison Formation
A stable isotopic investigation of resource partitioning among neosauropod dinosaurs of the Upper Jurassic Morrison Formation Benjamin T. Breeden, III SID: 110305422 [email protected] GEOL394H University of Maryland, College Park, Department of Geology 29 April 2011 Advisors: Dr. Thomas R. Holtz1, Jr., Dr. Alan Jay Kaufman1, and Dr. Matthew T. Carrano2 1: University of Maryland, College Park, Department of Geology 2: National Museum of Natural History, Department of Paleobiology ABSTRACT For more than a century, morphological studies have been used to attempt to understand the partitioning of resources in the Morrison Fauna, particularly between members of the two major clades of neosauropod (long-necked, megaherbivorous) dinosaurs: Diplodocidae and Macronaria. While it is generally accepted that most macronarians fed 3-5m above the ground, the feeding habits of diplodocids are somewhat more enigmatic; it is not clear whether diplodocids fed higher or lower than macronarians. While many studies exploring sauropod resource portioning have focused on differences in the morphologies of the two groups, few have utilized geochemical evidence. Stable isotope geochemistry has become an increasingly common and reliable means of investigating paleoecological questions, and due to the resistance of tooth enamel to diagenetic alteration, fossil teeth can provide invaluable paleoecological and behavioral data that would be otherwise unobtainable. Studies in the Ituri Rainforest in the Democratic Republic of the Congo, have shown that stable isotope ratios measured in the teeth of herbivores reflect the heights at which these animals fed in the forest due to isotopic variation in plants with height caused by differences in humidity at the forest floor and the top of the forest exposed to the atmosphere. -

Dr. Sahanaj Jamil Associate Professor of Botany M.L.S.M. College, Darbhanga
Subject BOTANY Paper No V Paper Code BOT521 Topic Taxonomy and Diversity of Seed Plant: Gymnosperms & Angiosperms Dr. Sahanaj Jamil Associate Professor of Botany M.L.S.M. College, Darbhanga BOTANY PG SEMESTER – II, PAPER –V BOT521: Taxonomy and Diversity of seed plants UNIT- I BOTANY PG SEMESTER – II, PAPER –V BOT521: Taxonomy and Diversity of seed plants Classification of Gymnosperms. # Robert Brown (1827) for the first time recognized Gymnosperm as a group distinct from angiosperm due to the presence of naked ovules. BENTHAM and HOOKSER (1862-1883) consider them equivalent to dicotyledons and monocotyledons and placed between these two groups of angiosperm. They recognized three classes of gymnosperm, Cyacadaceae, coniferac and gnetaceae. Later ENGLER (1889) created a group Gnikgoales to accommodate the genus giankgo. Van Tieghem (1898) treated Gymnosperm as one of the two subdivision of spermatophyte. To accommodate the fossil members three more classes- Pteridospermae, Cordaitales, and Bennettitales where created. Coulter and chamberlain (1919), Engler and Prantl (1926), Rendle (1926) and other considered Gymnosperm as a division of spermatophyta, Phanerogamia or Embryoptyta and they further divided them into seven orders: - i) Cycadofilicales ii) Cycadales iii) Bennettitales iv) Ginkgoales v) Coniferales vi) Corditales vii) Gnetales On the basis of wood structure steward (1919) divided Gymnosperm into two classes: - i) Manoxylic ii) Pycnoxylic The various classification of Gymnosperm proposed by various workers are as follows: - i) Sahni (1920): - He recognized two sub-divison in gymnosperm: - a) Phylospermae b) Stachyospermae BOTANY PG SEMESTER – II, PAPER –V BOT521: Taxonomy and Diversity of seed plants ii) Classification proposed by chamber lain (1934): - He divided Gymnosperm into two divisions: - a) Cycadophyta b) Coniterophyta iii) Classification proposed by Tippo (1942):- He considered Gymnosperm as a class of the sub- phylum pteropsida and divided them into two sub classes:- a) Cycadophyta b) Coniferophyta iv) D. -

Retallack 2021 Coal Balls
Palaeogeography, Palaeoclimatology, Palaeoecology 564 (2021) 110185 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Modern analogs reveal the origin of Carboniferous coal balls Gregory Retallack * Department of Earth Science, University of Oregon, Eugene, Oregon 97403-1272, USA ARTICLE INFO ABSTRACT Keywords: Coal balls are calcareous peats with cellular permineralization invaluable for understanding the anatomy of Coal ball Pennsylvanian and Permian fossil plants. Two distinct kinds of coal balls are here recognized in both Holocene Histosol and Pennsylvanian calcareous Histosols. Respirogenic calcite coal balls have arrays of calcite δ18O and δ13C like Carbon isotopes those of desert soil calcic horizons reflecting isotopic composition of CO2 gas from an aerobic microbiome. Permineralization Methanogenic calcite coal balls in contrast have invariant δ18O for a range of δ13C, and formed with anaerobic microbiomes in soil solutions with bicarbonate formed by methane oxidation and sugar fermentation. Respiro genic coal balls are described from Holocene peats in Eight Mile Creek South Australia, and noted from Carboniferous coals near Penistone, Yorkshire. Methanogenic coal balls are described from Carboniferous coals at Berryville (Illinois) and Steubenville (Ohio), Paleocene lignites of Sutton (Alaska), Eocene lignites of Axel Heiberg Island (Nunavut), Pleistocene peats of Konya (Turkey), and Holocene peats of Gramigne di Bando (Italy). Soils and paleosols with coal balls are neither common nor extinct, but were formed by two distinct soil microbiomes. 1. Introduction and Royer, 2019). Although best known from Euramerican coal mea sures of Pennsylvanian age (Greb et al., 1999; Raymond et al., 2012, Coal balls were best defined by Seward (1895, p. -

Géologique Nord Annales
SOCIÉTÉ GÉOLOGIQUE D U NORD ANNALES Tome CV 1985 3me trimestre IRIS - LILLIAD - Université Lille 1 SOCIETE GEOLOGIQUE DU NORD Extraits des Statuts Article 2. — Cette Société a pour objet de concourir à l'avancement de la géologie en général, et particulièrement de la géologie de la région du Nord de la France. — La Société se réunit de droit une fols par mois, sauf pendant la période des vacances. Elle peut tenir des séances extraordinaires décidées par le Conseil d'Administration. — La Société publie des Annale3 et des Mémoires. Ces publications sont mises en vente selon un tarif établi par le Conseil. Les Sociétaires bénéficient d'un tarif préférentiel f.1). Article 5. Le nombre des membres de la Société est Illimité. Pour faire partie de la Société, il faut s'être fait présenter dans l'une de ses séances par deux membres de la Société qui auront signé la présentation, et avoir été proclamé membre au cours de la séance suivante. Extraits du Règlement Intérieur § 7. — Les Annales et leur supplément constituent le compte rendu des séances. § 13. — Seuls les membres ayant acquitté leurs cotisation et abonnement de l'année peuvent publier dans les Annales. L'ensemble des notes présentées au cours d'une même année, par un même auteur, ne peut dépasser le total de 10 pages, 1 planche simili étant comptée pour 2 p. 1/2 de texte. Le Conseil peut, par décision spéciale, autoriser la publication de notes plus longues. § 17. — Les notes et mémoires originaux (texte et illustration) communiqués à la Société et destinés eux Annales doivent être remis au Secrétariat le jour même de leur présentation. -

Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation
i1 Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation William A. DiMichele, William E. Stein, and Richard M. Bateman DiMichele, W.A., Stein, W.E., and Bateman, R.M. 2001. Ecological sorting of vascular plant classes during the Paleozoic evolutionary radiation. In: W.D. Allmon and D.J. Bottjer, eds. Evolutionary Paleoecology: The Ecological Context of Macroevolutionary Change. Columbia University Press, New York. pp. 285-335 THE DISTINCTIVE BODY PLANS of vascular plants (lycopsids, ferns, sphenopsids, seed plants), corresponding roughly to traditional Linnean classes, originated in a radiation that began in the late Middle Devonian and ended in the Early Carboniferous. This relatively brief radiation followed a long period in the Silurian and Early Devonian during wrhich morphological complexity accrued slowly and preceded evolutionary diversifications con- fined within major body-plan themes during the Carboniferous. During the Middle Devonian-Early Carboniferous morphological radiation, the major class-level clades also became differentiated ecologically: Lycopsids were cen- tered in wetlands, seed plants in terra firma environments, sphenopsids in aggradational habitats, and ferns in disturbed environments. The strong con- gruence of phylogenetic pattern, morphological differentiation, and clade- level ecological distributions characterizes plant ecological and evolutionary dynamics throughout much of the late Paleozoic. In this study, we explore the phylogenetic relationships and realized ecomorphospace of reconstructed whole plants (or composite whole plants), representing each of the major body-plan clades, and examine the degree of overlap of these patterns with each other and with patterns of environmental distribution. We conclude that 285 286 EVOLUTIONARY PALEOECOLOGY ecological incumbency was a major factor circumscribing and channeling the course of early diversification events: events that profoundly affected the structure and composition of modern plant communities. -

The Joggins Fossil Cliffs UNESCO World Heritage Site: a Review of Recent Research
The Joggins Fossil Cliffs UNESCO World Heritage site: a review of recent research Melissa Grey¹,²* and Zoe V. Finkel² 1. Joggins Fossil Institute, 100 Main St. Joggins, Nova Scotia B0L 1A0, Canada 2. Environmental Science Program, Mount Allison University, Sackville, New Brunswick E4L 1G7, Canada *Corresponding author: <[email protected]> Date received: 28 July 2010 ¶ Date accepted 25 May 2011 ABSTRACT The Joggins Fossil Cliffs UNESCO World Heritage Site is a Carboniferous coastal section along the shores of the Cumberland Basin, an extension of Chignecto Bay, itself an arm of the Bay of Fundy, with excellent preservation of biota preserved in their environmental context. The Cliffs provide insight into the Late Carboniferous (Pennsylvanian) world, the most important interval in Earth’s past for the formation of coal. The site has had a long history of scientific research and, while there have been well over 100 publications in over 150 years of research at the Cliffs, discoveries continue and critical questions remain. Recent research (post-1950) falls under one of three categories: general geol- ogy; paleobiology; and paleoenvironmental reconstruction, and provides a context for future work at the site. While recent research has made large strides in our understanding of the Late Carboniferous, many questions remain to be studied and resolved, and interest in addressing these issues is clearly not waning. Within the World Heritage Site, we suggest that the uppermost formations (Springhill Mines and Ragged Reef), paleosols, floral and trace fossil tax- onomy, and microevolutionary patterns are among the most promising areas for future study. RÉSUMÉ Le site du patrimoine mondial de l’UNESCO des falaises fossilifères de Joggins est situé sur une partie du littoral qui date du Carbonifère, sur les rives du bassin de Cumberland, qui est une prolongation de la baie de Chignecto, elle-même un bras de la baie de Fundy. -
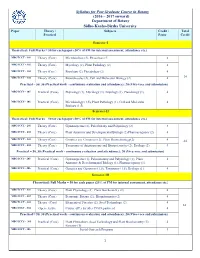
Syllabus for Post Graduate Course in Botany (2016 – 2017 Onward)
Syllabus for Post Graduate Course in Botany (2016 – 2017 onward) Department of Botany Sidho-Kanho-Birsha University Paper Theory / Subjects Credit / Total Practical Paper Credit Semester-I Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 101 Theory (Core) Microbiology (2), Phycology (2) 4 MBOTCCT - 102 Theory (Core) Mycology (2), Plant Pathology (2) 4 MBOTCCT - 103 Theory (Core) Bryology (2), Pteridology (2) 4 MBOTCCT - 104 Theory (Core) Biomolecules (2), Cell and Molecular Biology (2) 4 24 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 105 Practical (Core) Phycology (1), Mycology (1), Bryology (1), Pteridology (1). 4 MBOTCCS - 106 Practical (Core) Microbiology (1.5), Plant Pathology (1), Cell and Molecular 4 Biology (1.5). Semester-II Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 201 Theory (Core) Gymnosperms (2), Paleobotany and Palynology (2) 4 MBOTCCT - 202 Theory (Core) Plant Anatomy and Developmental Biology (2) Pharmacognosy (2) 4 MBOTCCT - 203 Theory (Core) Genetics and Genomics (2), Plant Biotechnology(2) 4 24 MBOTCCT - 204 Theory (Core) Taxonomy of Angiosperms and Biosystematics (2), Ecology (2) 4 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 205 Practical (Core) Gymnosperms (1), Palaeobotany and Palynology (1), Plant 4 Anatomy & Developmental Biology (1), Pharmacognosy (1). MBOTCCS - 206 Practical (Core) Genetics and Genomics (1.5), Taxonomy (1.5), Ecology (1). 4 Semester-III Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 301 Theory (Core) Plant Physiology (2), Plant Biochemistry (2) 4 MBOTCCT - 302 Theory (Core) Economic Botany (2), Bioinformatics (2) 4 MBOTCCT - 303 Theory (Core) Elements of Forestry (2), Seed Technology (2). -

A Reappraisal of Mississippian (Tournaisian and Visean) Adpression Floras from Central and Northwestern Europe
Zitteliana A 54 (2014) 39 A reappraisal of Mississippian (Tournaisian and Visean) adpression floras from central and northwestern Europe Maren Hübers1, Benjamin Bomfleur2*, Michael Krings3, Christian Pott2 & Hans Kerp1 1Forschungsstelle für Paläobotanik am Geologisch-Paläontologischen Institut, Westfälische Zitteliana A 54, 39 – 52 Wilhelms-Universität Münster, Heisenbergstraße 2, 48149 Münster, Germany München, 31.12.2014 2Swedish Museum of Natural History, Department of Paleobiology, Box 50007, SE-104 05, Stockholm, Sweden Manuscript received 3Department für Geo- und Umweltwissenschaften, Paläontologie und Geobiologie, Ludwig- 01.02.2014; revision Maximilians-Universität, and Bayerische Staatssammlung für Paläontologie und Geologie, Richard- accepted 07.04.2014 Wagner-Straße 10, 80333 Munich, Germany ISSN 1612 - 412X *Author for correspondence and reprint requests: E-mail: [email protected] Abstract Mississippian plant fossils are generally rare, and in central and northwestern Europe especially Tournaisian to middle Visean fossil floras are restricted to isolated occurrences. While sphenophytes and lycophytes generally are represented by only a few widespread and long-ranging taxa such as Archaeocalamites radiatus, Sphenophyllum tenerrimum and several species of Lepidodendropsis and Lepido- dendron, Visean floras in particular show a remarkably high diversity of fern-like foliage, including filiform types (Rhodea, Diplotmema), forms with bipartite fronds (Sphenopteridium, Diplopteridium, Spathulopteris, Archaeopteridium), others with monopodial, pinnate fronds (Anisopteris, Fryopsis) and still others characterized by several-times pinnate fronds (e.g., Adiantites, Triphyllopteris, Sphenopteris, Neu- ropteris). Most of these leaf types have been interpreted as belonging to early seed ferns, whereas true ferns seem to have been rare or lacking in impression/compression floras. In the upper Visean, two types of plant assemblages can be distinguished, i.e., the northern Kohlenkalk-type and the south-eastern Kulm-type assemblage. -

The Upper Carboniferous-Lower Permian Flora of Zöbing, Lower Austria
The upper Carboniferous-Lower Permian flora of Zöbing, Lower Austria Utrecht University Master Thesis Earth, Life and Climate By Koen Paalman 3470423 [email protected] 1 Contents page Abstract 3 1. Introduction 4 2. Geography, Geology and Lithology 6 3. Vegetation types 8 4. Different taxonomic groups 8 4.1 Calamites and other sphenopsids 8 4.2 Tree ferns 8 4.3 Cordaites 8 4.4 Pteridosperms 9 4.5 The medullosan pteridosperms 9 4.6 The peltaspermalean pteridosperms 9 5. Methods 10 6. Results 10 7. Correlation, comparison and interpretation 11 8. Discussion and conclusion 13 9. Acknowledgements 13 10. Appendix 14 10.1 Species list 14 10.2 Reference 16 11. Plates 19 2 Abstract During the Late Carboniferous and early Permian, a major floristic change took place in Euramerica. Gymnosperms replaced the previously dominant pteridophytes. This reflects a climatic change, i.e. from wetland-dominated to more arid conditions. Extensive studies on the vegetation during this time interval have recently been carried out in the Czech Republic. The Zöbing formation in Austria, is of the same age of these Czech formations, but has not yet been compared to them. Material from the Zöbing formation has been examined and compared to Czech floras. A clear transition can be seen in the flora of the Zöbing formation, from a Stephanian tree fern and Alethopteris zeilleri dominated flora, to a Asselian flora dominated by peltasperms and conifers. There are clear similarities between the floras of the Zöbing formation and the different Czechian formations, despite notable differences in species composition and abundance between Zöbing and the Czech formations. -

The Joggins Cliffs of Nova Scotia: B2 the Joggins Cliffs of Nova Scotia: Lyell & Co's "Coal Age Galapagos" J.H
GAC-MAC-CSPG-CSSS Pre-conference Field Trips A1 Contamination in the South Mountain Batholith and Port Mouton Pluton, southern Nova Scotia HALIFAX Building Bridges—across science, through time, around2005 the world D. Barrie Clarke and Saskia Erdmann A2 Salt tectonics and sedimentation in western Cape Breton Island, Nova Scotia Ian Davison and Chris Jauer A3 Glaciation and landscapes of the Halifax region, Nova Scotia Ralph Stea and John Gosse A4 Structural geology and vein arrays of lode gold deposits, Meguma terrane, Nova Scotia Rick Horne A5 Facies heterogeneity in lacustrine basins: the transtensional Moncton Basin (Mississippian) and extensional Fundy Basin (Triassic-Jurassic), New Brunswick and Nova Scotia David Keighley and David E. Brown A6 Geological setting of intrusion-related gold mineralization in southwestern New Brunswick Kathleen Thorne, Malcolm McLeod, Les Fyffe, and David Lentz A7 The Triassic-Jurassic faunal and floral transition in the Fundy Basin, Nova Scotia Paul Olsen, Jessica Whiteside, and Tim Fedak Post-conference Field Trips B1 Accretion of peri-Gondwanan terranes, northern mainland Nova Scotia Field Trip B2 and southern New Brunswick Sandra Barr, Susan Johnson, Brendan Murphy, Georgia Pe-Piper, David Piper, and Chris White The Joggins Cliffs of Nova Scotia: B2 The Joggins Cliffs of Nova Scotia: Lyell & Co's "Coal Age Galapagos" J.H. Calder, M.R. Gibling, and M.C. Rygel Lyell & Co's "Coal Age Galapagos” B3 Geology and volcanology of the Jurassic North Mountain Basalt, southern Nova Scotia Dan Kontak, Jarda Dostal,