The Case of Common Bean (Phaseolus Vulgaris L.)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
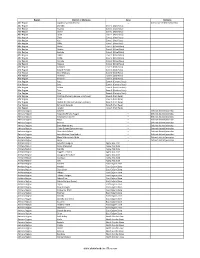
Districts of Ethiopia
Region District or Woredas Zone Remarks Afar Region Argobba Special Woreda -- Independent district/woredas Afar Region Afambo Zone 1 (Awsi Rasu) Afar Region Asayita Zone 1 (Awsi Rasu) Afar Region Chifra Zone 1 (Awsi Rasu) Afar Region Dubti Zone 1 (Awsi Rasu) Afar Region Elidar Zone 1 (Awsi Rasu) Afar Region Kori Zone 1 (Awsi Rasu) Afar Region Mille Zone 1 (Awsi Rasu) Afar Region Abala Zone 2 (Kilbet Rasu) Afar Region Afdera Zone 2 (Kilbet Rasu) Afar Region Berhale Zone 2 (Kilbet Rasu) Afar Region Dallol Zone 2 (Kilbet Rasu) Afar Region Erebti Zone 2 (Kilbet Rasu) Afar Region Koneba Zone 2 (Kilbet Rasu) Afar Region Megale Zone 2 (Kilbet Rasu) Afar Region Amibara Zone 3 (Gabi Rasu) Afar Region Awash Fentale Zone 3 (Gabi Rasu) Afar Region Bure Mudaytu Zone 3 (Gabi Rasu) Afar Region Dulecha Zone 3 (Gabi Rasu) Afar Region Gewane Zone 3 (Gabi Rasu) Afar Region Aura Zone 4 (Fantena Rasu) Afar Region Ewa Zone 4 (Fantena Rasu) Afar Region Gulina Zone 4 (Fantena Rasu) Afar Region Teru Zone 4 (Fantena Rasu) Afar Region Yalo Zone 4 (Fantena Rasu) Afar Region Dalifage (formerly known as Artuma) Zone 5 (Hari Rasu) Afar Region Dewe Zone 5 (Hari Rasu) Afar Region Hadele Ele (formerly known as Fursi) Zone 5 (Hari Rasu) Afar Region Simurobi Gele'alo Zone 5 (Hari Rasu) Afar Region Telalak Zone 5 (Hari Rasu) Amhara Region Achefer -- Defunct district/woredas Amhara Region Angolalla Terana Asagirt -- Defunct district/woredas Amhara Region Artuma Fursina Jile -- Defunct district/woredas Amhara Region Banja -- Defunct district/woredas Amhara Region Belessa -- -

Microstructural Differences Among Adzuki Bean (Vigna Angularis) Cultivars
Food Structure Volume 11 Number 2 Article 9 1992 Microstructural Differences Among Adzuki Bean (Vigna Angularis) Cultivars Anup Engquist Barry G. Swanson Follow this and additional works at: https://digitalcommons.usu.edu/foodmicrostructure Part of the Food Science Commons Recommended Citation Engquist, Anup and Swanson, Barry G. (1992) "Microstructural Differences Among Adzuki Bean (Vigna Angularis) Cultivars," Food Structure: Vol. 11 : No. 2 , Article 9. Available at: https://digitalcommons.usu.edu/foodmicrostructure/vol11/iss2/9 This Article is brought to you for free and open access by the Western Dairy Center at DigitalCommons@USU. It has been accepted for inclusion in Food Structure by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected]. FOOD STRUCTURE, Vol. II (1992), pp. 171-179 1046-705X/92$3.00+ .00 Scanning Microscopy International , Chicago (AMF O'Hare), IL 60666 USA MICROSTRUCTURAL DIFFERENCES AMONG ADZUKI BEAN (Vigna angularis) CULTIVARS An up Engquist and Barry G. Swanson Department of Food Science and Human Nutrition Washington State University, Pullman, WA 99164-6376 Abstract Introduction Scanning electron microscopy (SEM) was used to Adzuki beans are one of the oldest cultivated beans study mi crostructural differences among five adzuki bean in the Orient, often used for human food, prepared as a cultivars: Erimo, Express, Hatsune, Takara and VBSC. bean paste used in soups and confections (Tjahjadi and Seed coat surfaces showed different patterns of cracks , Breene, 1984). The starch content of adzuki beans is pits and deposits . Cross-sections of the seed coats re about 50 %, while the protein content ranges between vealed well organized layers of elongated palisade cell s 20%-25% (Tjahjadi and Breene, 1984) . -

The Diversity and Composition of Woody Plants in Chebera Churcura National Park (CCNP), Southern Ethiopia
Open Journal of Forestry, 2018, 8, 439-458 http://www.scirp.org/journal/ojf ISSN Online: 2163-0437 ISSN Print: 2163-0429 The Diversity and Composition of Woody Plants in Chebera Churcura National Park (CCNP), Southern Ethiopia Mezmir Girma1, Melesse Maryo2 1Shashemene Botanical Garden, Ethiopian Biodiversity Institute (EBI), Shashemene, Ethiopia 2Dilla University, Department of Biology (DU), Dilla, Ethiopia How to cite this paper: Girma, M., & Abstract Maryo, M. (2018). The Diversity and Com- position of Woody Plants in Chebera Chur- National parks harbor the most valuable biological diversities of national and cura National Park (CCNP), Southern Ethi- global significance. The study of these diversities assists for the proper re- opia. Open Journal of Forestry, 8, 439-458. source management. This study was carried out to determine woody species https://doi.org/10.4236/ojf.2018.84028 diversity and composition in CCNP. Systematic sampling was used to collect Received: January 2, 2018 vegetation data from 67 plots (400 m2 each), which were laid following altitu- Accepted: August 18, 2018 dinal gradient. From each plot presence/absence of woody species, abundance Published: August 21, 2018 and structural data were recorded. R Program, Sorensen’s similarity, Shannon Copyright © 2018 by authors and diversity index were used for vegetation analysis. Our result indicated that a Scientific Research Publishing Inc. total of 106 plant specimens belonging to 42 families and 90 genera were iden- This work is licensed under the Creative tified, of which 57.6% were trees, followed by 18.8% shrubs, 7.6% liana and Commons Attribution International License (CC BY 4.0). -

Seed Mineral Composition and Protein Content of Faba Beans (Vicia Faba L.) with Contrasting Tannin Contents
agronomy Article Seed Mineral Composition and Protein Content of Faba Beans (Vicia faba L.) with Contrasting Tannin Contents Hamid Khazaei * and Albert Vandenberg Department of Plant Sciences, University of Saskatchewan, Saskatoon, SK S7N 5A8, Canada; [email protected] * Correspondence: [email protected]; Tel.: +1-306-966-5859 Received: 17 March 2020; Accepted: 28 March 2020; Published: 3 April 2020 Abstract: Two-thirds of the world’s population are at risk of deficiency in one or more essential mineral elements. The high concentrations of essential mineral elements in pulse seeds are fundamentally important to human and animal nutrition. In this study, seeds of 25 genotypes of faba bean (12 low-tannin and 13 normal-tannin genotypes) were evaluated for mineral nutrients and protein content in three locations in Western Canada during 2016–2017. Seed mineral concentrations were examined by Inductively Coupled Plasma Mass Spectrometry (ICP-MS) and the protein content was determined by Near-Infrared (NIR) spectroscopy. Location and year (site-year) effects were significant for all studied minerals, with less effect for calcium (Ca) and protein content. Genotype by environment interactions were found to be small for magnesium (Mg), cobalt (Co), Ca, zinc (Zn), and protein content. Higher seed concentrations of Ca, manganese (Mn), Mg, and cadmium (Cd) were observed for low-tannin genotypes compared to tannin-containing genotypes. The protein content was 1.9% higher in low-tannin compared to tannin-containing genotypes. The high estimated heritability for concentrations of seed Mg, Ca, Mn, potassium (K), sulphur (S), and protein content in this species suggests that genetic improvement is possible for mineral elements. -

Phaseolus Vulgaris (Beans)
1 Phaseolus vulgaris (Beans) Phaseolus vulgaris (Beans) dry beans are Brazil, Mexico, China, and the USA. Annual production of green beans is around 4.5 P Gepts million tonnes, with the largest production around Copyright ß 2001 Academic Press the Mediterranean and in the USA. doi: 10.1006/rwgn.2001.1749 Common bean was used to derive important prin- ciples in genetics. Mendel used beans to confirm his Gepts, P results derived in peas. Johannsen used beans to illus- Department of Agronomy and Range Science, University trate the quantitative nature of the inheritance of cer- of California, Davis, CA 95616-8515, USA tain traits such as seed weight. Sax established the basic methodology to identify quantitative trait loci (for seed weight) via co-segregation with Mendelian mar- Beans usually refers to food legumes of the genus kers (seed color and color pattern). The cultivars of Phaseolus, family Leguminosae, subfamily Papilio- common bean stem from at least two different domes- noideae, tribe Phaseoleae, subtribe Phaseolinae. The tications, in the southern Andes and Mesoamerica. In genus Phaseolus contains some 50 wild-growing spe- turn, their respective wild progenitors in these two cies distributed only in the Americas (Asian Phaseolus regions have a common ancestor in Ecuador and have been reclassified as Vigna). These species repre- northern Peru. This knowledge of the evolution of sent a wide range of life histories (annual to perennial), common bean, combined with recent advances in the growth habits (bush to climbing), reproductive sys- study of the phylogeny of the genus, constitute one of tems, and adaptations (from cool to warm and dry the main current attractions of beans as genetic organ- to wet). -

Effects of Conservation-Induced Displacement on the Bacha of Southwest Ethiopia
Ethiopian Journal of the Social Sciences and Humanities (EJOSSAH): V. 14, No. 2 ISSN (online): 2520-582X ISSN (print): 1810-4487 Effects of Conservation-induced Displacement on the Bacha of Southwest Ethiopia Deresse Daniel† and Abeje Berhanu Abstract This paper examines the impact of conservation-induced displacement on the Bacha community resulting from the establishment of Chebera-Chuchura National Park in Konta Special Woreda, Southern Nations, Nationalities and Peoples Region (SNNPR). It attempts to reconstruct the customary functions that the forestland had played for the livelihoods of the Bacha community and its role in cementing reciprocal relations among different social groups in pre-eviction contexts. The study employed a combination of both qualitative and quantitative research tools including in-depth interviews, key informant interviews, focus group discussions, field observations and household survey. Michael Cernea’s analytical framework to assess the risks associated with displacement – Impoverishment Risk and Reconstruction (IRR) – is employed to see the multifaceted aspects of conservation-induced displacement. The findings revealed that in spite of the fact that displaced Bacha people have enjoyed greater level of access to land they suffered loss of entitlements to forest-based assets such as honey and plants of enormous medicinal value. The community also faced loss of job opportunities due to restrictions imposed on access to forestland that supported beneficial biodiversity for the livelihood of the Bacha people. Also adversely affected are inter-community relations built on reciprocal exchange of goods and services between the Bacha and neighboring farming communities. Now, the Bacha have faced the difficult task of adapting to the land-based crop farming as a new source of livelihood since they lack the necessary farming skills to make a living out of crop farming. -

Breeding Technology Assessment at Small Holder Dairy Cattle Production Level in Selected Districts of HYDYA ZONE, Southern Ethiopia
Advances in Life Science and Technology www.iiste.org ISSN 2224-7181 (Paper) ISSN 2225-062X (Online) Vol.34, 2015 Breeding Technology Assessment at Small Holder Dairy Cattle Production Level in Selected Districts of HYDYA ZONE, Southern Ethiopia Tsegay Lijalem 1, Mulatua Abebe 2 and Belachew Haile 2 1, Department of Animal and Range Sciences, Agriculture College, Wolaita Soddo University P.O. box,138, Wolaita, Ethiopia 2, Expert of Animal Production at Soro woreda, Hdiya Zone, Southern, Ethiopia Abstract The study was conducted from May to October 2014 in selected districts of Soro woreda, Hdiya Zone, Southern, Ethiopia. The objective of the study was to assess breeding technology on dairy cattle production at small holder level. To under taken the study, purposive sampling method was used. Data was obtained from both primary and secondary sources and the collected data were analyzed by using descriptive statistics. The study was found 398.76 average total livestock and 350.3 cattle per households in TLU. The study showed that only 39% of the respondents used artificial insemination for breeding their dairy cattle while 61% did not use. Respectively,42 %, 23%, 19% and 16% of respondents have said artificial insemination has advantage over natural mating because artificial insemination can use in accelerating introduction of new genetics, has low cost in comparison of bull price, no need of bull management and low disease transmission. On the other hand, due to heat period of cow could be easily detected by bull (66%) and no need of technicians support (34%) the respondents have said natural mating has advantage over artificial insemination. -

And Chick Pea (Cicer Arietinum L.) with Azospirillum Brasilense Strain Cd
Symbiosis, 21 (1996) 41-48 41 Balaban, Philadelphia/Rehovot Field Inoculation of Common Bean (Phaseolus vulgaris L.) and Chick Pea (Cicer arietinum L.) with Azospirillum brasilense Strain Cd SAUL BURDMAN, SHLOMO SARIC, JAIME KIGEL and YAACOV OKON* Faculty of Agriculture, The Hebrew University of Jerusalem, Rehovot 76100, Israel, Tel. +972-8-9481216, Fax. +972-8-466794 Received January 7, 1996; Accepted March 27, 1996 Abstract Field inoculation with Azospirillum brasilense strain Cd increased nodule dry weight (90%), plant-growth parameters and seed yield (99%) of naturally nodulated Cicer arieiinum L. (chick pea). In Phaseolus vulgaris L. (common bean), inoculation with Rhizobium etli TAL182 and R. tropici CIAT899 increased seed yield (13%), and combined inoculation with Rhizobium and Azospirillum resulted in a further increase (23%), while plants inoculated with Azospirillum alone did not differ in yield from uninoculated controls, despite a relative increase in shoot dry weight. Keywords: Phaseolus vulgaris L., Cicer arietinum L., Azospirillum, Rhizobium, nodulation 1. Introduction Data accumulated throughout the world over the past 20 years indicate that free-living rhizobacteria of the genus Azospirillum are capable of increasing the yield of important crops grown in various soils and climatic regions (Okon and Labandera-Gonzales, 1994). The plant growth-promoting effects of Azospirillum inoculation are attributed mainly to improved root development *rhe author to whom correspondence should be sent. 0334-5114/96/$05.50 ©1996 Balaban j 42 S. BURDMAN ET AL. and to the subsequent increase in the rate of water and mineral uptake. There is some evidence that the excretion of phytohormones by the bacteria may be responsible for the observed positive effects on root morphology and activity (Fallik et al., 1994). -

Army Worm Infestation in SNNP and Oromia Regions As of 24 May
Army worm infestation in SNNP and Oromia Regions As of 24 May, some 8,368 hectares of belg cropland was reportedly destroyed by army worms in Wolayita zone of SNNPR - an area that suffered from late onset of the 2013 belg rains and subsequent heavy rains that damaged belg crops. The damage caused by the army worms will further reduce the expected harvest this season. Similar incidents were also reported from Boricha, Bona Zuria, Dara, Dale, Hawassa Zuria and Loko Abaya woredas of Sidama zone; Loma and Mareka woredas of Dawro zone (SNNPR), as well as from drought prone areas of East and West Hararge zones of Oromia Region; and quickly spreading to neighbouring areas. In Boricha woreda, for example, more than 655 hectares of belg cropland was destroyed in the course of one week, this is indicative of the speed that damage is being caused. Immediate distribution of spraying containers and chemicals to the farmers is required to prevent further loss of belg crops. For more information, contact: [email protected] Health Update The number of meningitis cases has gradually declined since the outbreak was declared in January. To date, 1,371 cases were reported from 24 woredas in five zones of SNNP and Oromia Regions. The Government, with support from health partners, is conducting a reactive vaccination in the affected areas, with 1, 678,220 people vaccinated so far. Next week, the number of people vaccinated during the Addis Ababa City Administration meningitis vaccination campaign, conducted from 20 to 26 May, will be released. Meanwhile, the number of kebeles reporting cases of Yellow Fever in South Ari, Benatsemay and Selmago woredas of South Omo zone, SNNPR, increased. -

EFFECT of CHICKPEA (Cicer Arietinum
EFFECT OF CHICKPEA (Cicer arietinum L.) GERMINATION ON THE MAJOR GLOBULIN CONTENT AND IN VITRO DIGESTIBILITY1 Guilherme Vanucchi PORTARI2, Olga Luisa TAVANO2, Maraiza A. da SILVA2, Valdir Augusto NEVES2,* SUMMARY Chickpea seed germination was carried out over a period of 6 days. Little variation in the nitrogen and total globulin content was observed. The major globulin (11 S type) showed higher variation after the 4th day of germination. The elution behaviour and distribution of the isolated major globulin fraction on Sepharose CL-6B chromatography showed little modification at the end of germination. On SDS-PAGE the peak eluted from Sepharose CL-6B showed changes in protein bands between 20 and 30 kDa and above 60 kDa, indicating protein degradation during the period. Proteolytic activity was detected in the albumin fraction of the seeds, which increased up to the fourth and then decreased up to the sixth day, when isolated chickpea total globulin and casein were used as substrates. Chickpea flour, isolated albumin and total globulin fractions did not show an increase for in vitro digestibility; however, the isolated major globulin was more susceptible to hydrolysis after germination. Keywords: chickpea germination, protein fractions, major globulin, protease activity, in vitro digestibility. RESUMO EFEITO DA GERMINAÇÃO DE GRÃO-DE-BICO (Cicer arietinum L.) NA GLOBULINA MAJORITÁRIA E DIGESTIBILIDADE IN VITRO. A germinação das sementes de grão-de-bico foi acompanhada por um período de 6 dias, no qual pequenas variações nos teores de nitrogênio e globulina total foram registradas. A globulina majoritária (tipo 11 S) apresentou maiores variações após o quarto dia de germinação. -

World Bank Document
Sample Procurement Plan (Text in italic font is meant for instruction to staff and should be deleted in the final version of the PP) Public Disclosure Authorized (This is only a sample with the minimum content that is required to be included in the PAD. The detailed procurement plan is still mandatory for disclosure on the Bank’s website in accordance with the guidelines. The initial procurement plan will cover the first 18 months of the project and then updated annually or earlier as necessary). I. General 1. Bank’s approval Date of the procurement Plan: Updated Procurement Plan, M 2. Date of General Procurement Notice: Dec 24, 2006 Public Disclosure Authorized 3. Period covered by this procurement plan: The procurement period of project covered from year June 2010 to December 2012 II. Goods and Works and non-consulting services. 1. Prior Review Threshold: Procurement Decisions subject to Prior Review by the Bank as stated in Appendix 1 to the Guidelines for Procurement: [Thresholds for applicable procurement methods (not limited to the list below) will be determined by the Procurement Specialist /Procurement Accredited Staff based on the assessment of the implementing agency’s capacity.] Public Disclosure Authorized Procurement Method Prior Review Comments Threshold US$ 1. ICB and LIB (Goods) Above US$ 500,000 All 2. NCB (Goods) Above US$ 100,000 First contract 3. ICB (Works) Above US$ 15 million All 4. NCB (Works) Above US$ 5 million All 5. (Non-Consultant Services) Below US$ 100,000 First contract [Add other methods if necessary] 2. Prequalification. Bidders for _Not applicable_ shall be prequalified in accordance with the provisions of paragraphs 2.9 and 2.10 of the Public Disclosure Authorized Guidelines. -

Tsedeke Lambore Gemecho Hawassa University, Ethiopia +251-91-381 0141 Personal Email: [email protected] Institutional Email: [email protected]
Curriculum Vitae Tsedeke Lambore Gemecho Hawassa University, Ethiopia +251-91-381 0141 Personal Email: [email protected] Institutional Email: [email protected] Personal Information Full Name: Tsedeke Lambore Gemecho Date of Birth: Meskerem 16, 1972 E.C (September 27, 1979 G.C) Place of Birth: Duna Woreda, Hadiya Zone, SNNPR, Ethiopia Languages: English and Amharic (Excellent in spoken and written) University or College Training/ Qualifications PhD Student in Applied Statistics • Hawassa University (Starting November 2014) • PhD Thesis title: “ Generalized Linear Mixed Modelling of Determinants of International Migration from the Southern Ethiopia: Small Area Estimation Approach “, the proposal was presented in 5th ISBALO conference of African Young Statisticians which was held June 13-17, 2016 Pretoria, Republic of South Africa. • Also, PhD Thesis proposal was presented at Hawassa University, Ethiopia. • A paper published in website “http://www.sciencepublishinggroup.com/j/ajtas ” was presented in annual research conference of Wachemo University, Ethiopia in 2016. • PhD thesis consists of four independent papers entitles: ° Logistic Mixed Modelling of Determinants of International Migration from the Southern Ethiopia: Small Area Estimation Approach (Published in http://www.sciencepublishinggroup.com/j/ajtas ). ° Count Regression Modelling on Number of International Migrants in Households (To be submitted for peer-review journals) ° Generalized Estimating Equations Analysis of Correlated Binary International Migrant Destination Country Choice (To be submitted for peer-review journals) ° Nonlinear Principal Component Analysis on Likert Scaled Push‒Pull Factors of International Migration (To be submitted for peer-review journals) • PhD Thesis is submitted to the school of Mathematical & Statistical Sciences and it will be defended on June 03-04, 2018.