Taxonomic Revision of Sorbus Subgenus Aria Occurring in the Czech Republic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

JOURNAL of JOURNAL of BOTANY Sorbus Hugh-Mcallisteri, Nom. Nov
Thaiszia - J. Bot., Košice, 18: 65-67, 2008 THAISZIA http://www.bz.upjs.sk/thaiszia/index.html JOURNAL OF BOTANY Sorbus hugh-mcallisteri , nom. nov. for Sorbus apiculata MCALLISTER 2005, nom. illeg. VLASTIMIL MIKOLÁŠ Hanojská 4, SK- 040 13 Košice, Slovakia, e-mail: [email protected] Mikoláš V. (2008): Sorbus hugh-mcallisteri nom. nov. for Sorbus apiculata McAllister 2005, nom. illeg. – Thaiszia – J. Bot. 18: 65-67. – ISSN 1210-0420. Abstract: Sorbus apiculata MCALLISTER 2005 is a later homonym of Sorbus apiculata (KOVANDA ) MIKOLÁŠ . The new name Sorbus hugh- mcallisteri MIKOLÁŠ is applied for the species. The species belongs to the section Multijugae (T.T.Y U) MCALL . It is known from NW Yunnan, China. Sorbus apiculata (KOVANDA ) MIKOLÁŠ 2004 is a member of Sorbus danubialis agg. ( Sorbus subg. Aria ) with the occurrence in Bohemia, Czech Republic. Keywords: Sorbus , Sorbus apiculata , S. hugh-mcallisteri , Czech Republic, China HUGH MCALLISTER , the best specialist of the genus of Sorbus s.s., recently described 28 species in his monograph of the genus (MCALLISTER 2005). The author accepts the genus Sorbus in its strict sense (with the type species S. acucuparia L)., splitting it up into 2 subgenera: subgenus Sorbus and subgenus Albocarmesinae MCALL . The latter includes species with fruits lacking chromoplasts and with white to crimson coloration. Some species have crimson petals, the character parallel to the European subg. Chamaemespilus (MEDICUS ) K.K OCH of the genus Sorbus (or Aria (PERSOON ) HOST if we accept the narrow concept of the genus Sorbus used by MCALLISTER ). The species of the subgenus are distributed in the Sino-Himalayan floristic region. -

Sehr Geehrte Damen Und Herren! Werte Mitbürgerinnen Und Mitbürger! Bei Der Gemeinderatssitzung Am 31
Sehr geehrte Damen und Herren! Werte Mitbürgerinnen und Mitbürger! Bei der Gemeinderatssitzung am 31. Okto in einem eigenen Termin am Freitag, den 14. ber 2013 wurden entscheidende Weichen für Februar 2014, um 19 Uhr im GH Holzer in die Zukunft unserer Gemeinde gestellt. formieren. Dabei wird auch ausgelotet, inwie Nach längeren Verhandlungen konnte der weit tatsächlich Interesse von Jugendlichen Vertrag mit dem Wohnbauträger „Austria“ unserer Gemeinde besteht – dies ist dann für abgeschlossen werden, und somit wird das den Wohnbauträger die Voraussetzung, ob „Betreute Wohnen“ bald Realität in unserer das Vorhaben realisiert wird. Gemeinde. Über die Wintermonate sollen Ich kann nur alle Jugendlichen, die gerne dann noch alle baurechtlichen Fakten abge ihren Start ins Leben in unserer Marktge schlossen werden – im Frühjahr wird gebaut. meinde Wullersdorf beginnen wollen, herzlich Ebenso kauften wir bei dieser Sitzung das einladen, von diesem Angebot Gebrauch zu restliche Areal von Herrn DI Daniel Brabenetz, machen! und beschlossen gleichzeitig auch einen Ver In diesem Sinne danke ich allen aktiven trag mit dem Wohnbauträger „Waldviertel“, Kräften in unserer Gemeinde für die stets welcher uns auch ein so genanntes „Junges gute Zusammenarbeit und wünsche Ihnen Wohnen“ ermöglichen soll. Diese Wohnan zum bevorstehenden Weihnachtsfest und lage würde dann unter anderem auf diesem dem Jahreswechsel alles erdenklich Gute, Areal errichtet und soll als „Startwohnungen“ vor allem Gesundheit. dienen. Ihr Richard HOGL e.h. Über das genaue Prozedere, die Voraus Bürgermeister setzungen und Bestimmungen werden wir (0676) 401 42 67 Der Dorftrommler Dezember 2013 Finanzielle Unterstützungen Insgesamt erhielt unsere Gemeinde hier mit im letzten Quartal 2013 Förderungen in durch das Land Niederösterreich der Höhe von R 48.338,65, plus einer Zusa R Die NÖ Landesregierung hat in Ihrer Sit ge über 35.000,–, wofür ich als Bürgermei zung vom 1. -

Samara Newsletter July & August 2020
SamaraThe International Newsletter of the Millennium Seed Bank Partnership Special issue featuring projects and research from The Global Tree Seed Bank Programme, funded by the Garfield Weston Foundation August/September 2020 Issue 35 ISSN 1475-8245 Juglans pyriformis in the State of Veracruz Conserving and investigating native tree seeds to support community-based reforestation initiatives in Mexico Veracruz Pronatura Photo: Mexico is the fourth richest country in the world in terms of plant Millennium Seed Bank. Seed research has species diversity, after Brazil, China, and Colombia with a flora of been carried out on 314 species to study ca. 23,000 vascular plants. Around half of the plant species are their tolerance to desiccation for seed endemic and nearly 3,500 are trees. banking and to determine germination requirements to inform propagation activities. One of the key project species ELENA CASTILLO-LORENZO (Latin America Projects Coordinator, RBG Kew), MICHAEL WAY is Cedrela odorata (Spanish cedar), whose (Conservation Partnership Coordinator (Americas, RBG Kew) & TIZIANA ULIAN (Senior Research conservation status is vulnerable (IUCN Leader – Diversity and Livelihoods, RBG Kew) 2020) due to exploitation for its highly Trees and forests provide multiple goods Iztacala of the Universidad Autónoma valued wood. C. odorata is also used for and benefits for humans, such as high- de México (Fes-I UNAM). The aim medicinal purposes by local communities quality wood, fruit, honey, and other of this project was to conserve tree in Mexico, with the leaves being prepared ecosystem services, including clean water, species through a collaborative research in herbal tea to treat toothache, earache, prevention of soil erosion and mitigation of programme focusing on endemic, and intestinal infections. -

Mittelalter.Pdf
Herrschaftsbildung und wirtschaftlicher Mißerfolg – Guntersdorf im späten Mittelalter (1258‐1476) Von Markus Jeitler Einleitung Die Geschichte unseres Ortes läßt sich in den Jahrzehnten nach seiner ersten 1 schriftlichen Nennung (1108 bzw. 1141)TPD DPT mangels erhaltener Quellen so gut wie nicht erfassen. Dies liegt vermutlich daran, daß ein großer Teil des Melker Stiftarchivs im Jahre 1297 einer Feuersbrunst zum Opfer fiel und somit wohl auch für unsere Fragestellungen relevante Stücke verloren gingen. Aus späteren Hinweisen und Überlieferungen kann man aber feststellen, daß das Stift Melk bis 1538 weiterhin die Lehenshoheit in Guntersdorf innehatte und seine Güter jeweils an Lehensträger ausgab, was noch auszuführen sein wird. An bisherigen Zusammenfassungen zur mittelalterlichen und neuzeitlichen Geschichte Guntersdorfs seien ein Beitrag von 2 Joseph Feil,TPD DPT dem der entsprechende Artikel in der „Topographie von 3 Niederösterreich“TPD DPT im wesentlichen folgt, und eine Zusammenstellung von Gertrude 4 Langer‐Ostrawsky im Heimatbuch des Bezirkes Hollabrunn genannt.TPD DPT Die Zeit der Übernahme der Herrschaft durch die Familie der Wallseer wird schließlich durch 5 Karel Hruza exemplarisch behandelt;TPD DPT Ignaz Franz Keiblinger geht in seinem umfangreichen Werk über die Geschichte des Stiftes Melk im Rahmen der 6 Darstellung der Pfarre Wullersdorf ebenfalls auf Guntersdorf ein.TPD DPT Darüber hinaus konnten weitere Hinweise gefunden werden, die das Gesamtbild abrunden bzw. für 7 Großnondorf neue Erkenntnisse bringen werden.TPD DPT Besitzungen zu Guntersdorf. Die ersten Nachrichten, die die Geschichte unseres Ortes im 13. Jahrhundert beleuchten, betreffen das Stift Heiligenkreuz. König Ottokar II. Přemysl bestätigt im Jahre 1258 die Schenkung des Albero von Leis über vier Mansen, aus seinem 8 Eigengut, die er nun zu seinem Seelenheil dem Stift übergibt.TPD DPT Diese Schenkung wird am 24. -

Klima- Und Energie-Modellregionen
Klima- und Energie-Modellregion : KLIMA- UND ENERGIEMODELLREGION PULKAUTAL Bericht der Umsetzungsphase Weiterführungsphase I Weiterführungsphase II Weiterführungsphase III Zwischenbericht Endbericht Inhaltsverzeichnis: 1. Fact-Sheet zur Klima- und Energie-Modellregion 2. Zielsetzung 3. Eingebundene Akteursgruppen 4. Aktivitätenbericht 5. Best Practice Beispiel der Umsetzung 1. Fact-Sheet zur Klima- und Energie-Modellregion Facts zur Klima- und Energie-Modellregion Name der Klima- und Energiemodellregion (KEM): Klima- und Energiemodellregion Pulkautal (Offizielle Regionsbezeichnung) Geschäftszahl der KEM B287567 Trägerorganisation, Rechtsform Initiative Pulkautal – Verein zur Entwicklungsförde- rung der Gemeinden des Gerichtbezirkes Haugsdorf Deckt sich die Abgrenzung und Bezeichnung der X Ja KEM mit einem bereits etablierten Regionsbegriff (j/n)? Initiative Pulkautal Falls ja, bitte Regionsbezeichnung anführen: Facts zur Klima- und Energiemodellregion: - Anzahl der Gemeinden: Die KEM umfasst insgesamt 6 Gemeinden - Anzahl der Einwohner/innen: 6.543 Einwohnern in der KEM - geografische Beschreibung (max. 400 Zeichen) Die Region Pulkautal liegt im nördlichen Weinviertel in Niederösterreich im politischen Bezirk Hol- labrunn. Das Pulkautal grenzt direkt an die Tsche- chische Republik. Namensgeber des Tales ist der Fluss Pulkau. Die Region liegt zwischen zwei Bal-lungszentren, ca. 80 Kilometer nord-westlich von Wien an der Grenze zu Tschechien, nur 15 Kilometer von Znaim und ca. 80 Kilometer von Brünn entfernt. Modellregions-Manager/in (MRM) -

Conifer Quarterly
Conifer Quarterly Vol. 24 No. 4 Fall 2007 Picea pungens ‘The Blues’ 2008 Collectors Conifer of the Year Full-size Selection Photo Credit: Courtesy of Stanley & Sons Nursery, Inc. CQ_FALL07_FINAL.qxp:CQ 10/16/07 1:45 PM Page 1 The Conifer Quarterly is the publication of the American Conifer Society Contents 6 Competitors for the Dwarf Alberta Spruce by Clark D. West 10 The Florida Torreya and the Atlanta Botanical Garden by David Ruland 16 A Journey to See Cathaya argyrophylla by William A. McNamara 19 A California Conifer Conundrum by Tim Thibault 24 Collectors Conifer of the Year 29 Paul Halladin Receives the ACS Annual Award of Merits 30 Maud Henne Receives the Marvin and Emelie Snyder Award of Merit 31 In Search of Abies nebrodensis by Daniel Luscombe 38 Watch Out for that Tree! by Bruce Appeldoorn 43 Andrew Pulte awarded 2007 ACS $1,000 Scholarship by Gerald P. Kral Conifer Society Voices 2 President’s Message 4 Editor’s Memo 8 ACS 2008 National Meeting 26 History of the American Conifer Society – Part One 34 2007 National Meeting 42 Letters to the Editor 44 Book Reviews 46 ACS Regional News Vol. 24 No. 4 CONIFER QUARTERLY 1 CQ_FALL07_FINAL.qxp:CQ 10/16/07 1:45 PM Page 2 PRESIDENT’S MESSAGE Conifer s I start this letter, we are headed into Afall. In my years of gardening, this has been the most memorable year ever. It started Quarterly with an unusually warm February and March, followed by the record freeze in Fall 2007 Volume 24, No 4 April, and we just broke a record for the number of consecutive days in triple digits. -

De Sorbus-Collectie in De Botanische Tuinen En Het Belmonte Arboretum Van De Landbouwhogeschool Te Wageningen
DRS. K. J. W. HENSEN DE SORBUS-COLLECTIE IN DE BOTANISCHE TUINEN EN HET BELMONTE ARBORETUM VAN DE LANDBOUWHOGESCHOOL TE WAGENINGEN IV (The Sorbits collection in the Botanical Gardens and Belmonte Arboretum of the Agricultural University at Wageningen. IV) In het 20ste Jaarboek van de Nederlandse Dendrologische Vereniging is een overzicht gegeven van de te Wageningen gekweekte Sorbus-pianten (HENSEN, 1957). Aanvullingen en wijzigingen verschenen in volgende jaarboeken (HENSEN, 1959 en 1963). Sedert de tekst van het laatste artikel geschreven werd, heeft opnieuw een aantal planten voor het eerst gebloeid en vrucht gedragen, zodat determinatie mogelijk werd. Van enige andere, reeds eerder in deze publikaties opgenomen planten werd de determinatie herzien. Deze aanvul lingen en wijzigingen van de vorige publikaties worden in dit artikel gepubliceerd. Aan gezien niet alle lezers van "Dendroflora" ook de Jaarboeken van de Nederlandse Dendrolo gische Vereniging ter beschikking zullen hebben, wordt aan het einde van dit artikel een lijst gegeven van alle taxa r), waarvan gedetermineerde planten in de Wageningense col lectie aanwezig zijn. De namen van soorten, variëteiten, apomicten of in het wild voorkomende hybriden worden gevolgd door de auteursnaam en een verwijzing naar de eerste publikatie van de naam. Deze wijziging bestaat uit de afgekorte titel, ev. het nummer van een boekdeel of jaargang van een tijdschrift, daarna de bladzijde en tenslotte het jaar van verschijnen. Namen van cultivars worden daarentegen gevolgd door de naam van de winner. Is deze niet bekend, dan laten wij de naam van de auteur volgen met een verwijzing naar de eerste publikatie van de naam. Synoniemen (cursief gedrukt) zijn slechts opgenomen, voor zover deze in botanische tuinen of in Nederlandse kwekerijen in gebruik zijn. -

A Quick Approach to Determine the Optimal Moisture Content For
AARHUS UNIVERSITET An approach to determine the optimal moisture content for controlled dormancy breakage in new tree and shrub species. Martin Jensen Department of Food Science Aarhus University [email protected] Background • physiological seed dormancy is released after a period of moist chilling • chilling at slightly reduced and controlled moisture content gives: - control of premature germination - ability to prolong the chilling duration - all seeds released from dormancy - ability to germinate at high or low temperatures and fast germination - surface dry seeds with less fungus attack - potential of retaining desiccation tolerance during entire chilling - ’advantage from more precise control’ Applied techniques change – from manual uncontrolled to mechanised controlled prototype to advanced controlled seed treatment CMC chilling Species MC %(f.w.) Duration Acer palmatum 35 – 37 8-12 w Acer platanoides 36 – 40 16-20 Acer pseudoplatanus 44 – 46 16-20 Amelanchier lamarckii 43 – 45 16-24 Berberis thunbergii 38 – 42 12-16 Fagus sylvatica 30 – 32 16-20 Fraxinus excelsior 42 – 44 16-20 Prunus avium 27 – 29 12-16 Quercus rubra 38 – 45 10-14 Sorbus aucuparia 43 – 45 16-20 Syringa vulgaris 45 8-16 Tilia cordata 40 – 43 16-24 Abies nordmanniana 32 – 34 8-10 Abies procera 30 – 34 8-10 Picea sitchensis 27 – 30 12-18 Pinus contorta 35 12-18 Pseudotsuga menziesii 32 – 35 12-18 Traditional investigation: - 5-8 moisture contents - 4-6 chilling durations - full factorial experiments - germination at 2 temperatures - several seed lots/ provenances = precise results, high costs, and long time experiments Problem in expanding the species list - Less economical important species - Some scientific and funding resistance of investigating a known principle on yet another species How can we go from 15 to 100 species with less economical input? Hypothesis: The optimal moisture content for controlled breakage of seed dormancy is always at or just below the critical moisture content for germination. -

SBCO-Bull41-P295-412
SESSION ALSACE - VOSGES - FORÊT NOIRE 295 39es Sessions extraordinaires - 2009 29 mai au 5 juin 10 au 17 juillet 2009 Alsace, Vosges et Forêt Noire Organisation scientifique et direction des excursions : Jean-Pierre BERCHTOLD Bulletin de la Société Botanique du Centre-Ouest - Nouvelle Série - Tome 41 - 2010 296 Y. PEYTOUREAU Les sessions de la Société Botanique du Centre-Ouest 1 1974 : Montendre (Charente-Maritime) 2 1975 : Nontron (Dordogne) 3 1976 : Mijanès (Ariège) 4 1977 : Jura 5 1978 : Saint-Junien (Haute-Vienne) 6 1979 : Corrèze 7 1980 : Cantal 8 1981 : Provence occidentale 9 1982 : Causses 10 1983 : Vosges et Alsace 11 1984 : Corse (session 11 bis en 1985) 12 1985 : Limousin 13 1986 : Causse-Comtal, Aubrac et Margeride 14 1987 : Haute-Cerdagne et Capcir 15 1988 : Haute-Normandie 16 1989 : Haute-Savoie 17 1990 : Littoral roussillonnais et audois 18 1991 : Queyras 19 1992 : Sud-Marocain 20 1992 : Marges nord-est de l’Île-de-France 21 1993 : Finistère 22 1994 : Nord - Pas-de-Calais 23 1995 : Charente-Maritime 24 1996 : Morbihan 25 1997 : Sud-est du Bassin Parisien 26 1998 : Hauts Cantons de l’Hérault et Larzac sud 27 1999 : Haut-Verdon 28 2000 : Partie orientale des Pyrénées 29 2001 : Vendée 30 2001 : Ténérife 31 2002 : Cotentin 32 2003 : Nord de la Corse 33 2004 : Provence calcaire et siliceuse 34 2005 : Haute-Marne et Côte-d’Or 35 2006 : Millau - Grands Causses 36 2007 : Jaca - Navarre/Aragon 37 2008 : Ubaye 38 2009 : Alsace, Vosges et Forêt Noire 39 2010 : Aude 40 2010 : Corse Photo au dos : Iris sibirica. Meistratzhiem (cliché Yann KELEN). -

Apró Közlemények / Short Communications
http://kitaibelia.unideb.hu/ ISSN 2064-4507 (Online) ● ISSN 1219-9672 (Print) © Department of Botany, University of Debrecen, Hungary 23 (1): 103–105.; 2018 DOI: 10.17542/kit.23.103 Apró közlemények / Short communications 1. Thelypteris palustris Schott a Börzsöny hegységben / Thelypteris palustris Schott in Börzsöny Mts (N Hungary) 2017. május 20-án a védett Thelypteris palustris Schott ismeretlen állományát fedezte fel Ferenczi Balázs a hegység forrásainak térképezése során Nagybörzsöny határában, a Lóhegyi- patak völgyében (N 47.89741°, E 18.90516°, tszf.: 425 m; KEF: 8179/1). A Börzsöny hegységből a tőzegpáfrányt NAGY (2007) nem említi. A szomszédos Ipoly- völgyből (SCHMOTZER 2008) és a Közép-Duna-völgyből ismertek előfordulásai, ugyanakkor középhegységeinkben ritka (BARTHA et al. 2015). 2017. november 1-én közös terepbejáráson mértük fel a populációt és a termőhelyéül szolgáló lápszemet. A gyertyános kocsánytalan tölgyesbe ágyazódó lápszem a délkeleti irányú völgy bal oldalán, egy, a talajból fakadó forrásnál helyezkedik el, feltételezhetően természetes eredetű vízállásos mélyedésben. Kiterjedése mintegy 300 négyzetméter (25 m × 10–15 m). Területének közel harmadát 1–8 m magas cserjék borítják (Cornus sanguinea, Salix cinerea, Sambucus nigra, Rubus fruticosus agg., Viburnum opulus). A Thelypteris palustris állomány összesen mintegy 40–50 négyzetmétert borít. A lápszem gyepszintje fejlett. A benne megfigyelt fajok a következők voltak: Aegopodium podagraria, Athyrium filix-femina, Callitriche sp., Caltha palustris subsp. laeta, Carex -
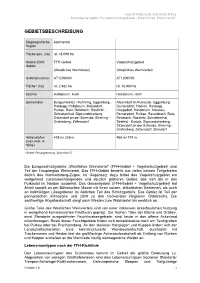
Gebietsbeschreibung
HAUPTREGION WEINVIERTEL Managementplan Europaschutzgebiete „Westliches Weinviertel“ GEBIETSBESCHREIBUNG Biogeografische kontinental Region Fläche ges. (ha) rd. 18.099 ha Natura 2000- FFH-Gebiet Vogelschutzgebiet Gebiet (Westliches Weinviertel) (Westliches Weinviertel) Gebietsnummer AT1209A00 AT1209V00 Fläche* (ha) rd. 2.982 ha rd. 16.904 ha Bezirke Hollabrunn, Horn Hollabrunn, Horn Gemeinden Burgschleinitz - Kühnring, Eggenburg, Alberndorf im Pulkautal, Eggenburg, Hardegg, Hollabrunn, Meiseldorf, Guntersdorf, Hadres, Hardegg, Pulkau, Retz, Retzbach, Röschitz, Haugsdorf, Hollabrunn, Maissau, Schrattenthal, Sigmundsherberg, Pernersdorf, Pulkau, Ravelsbach, Retz, Sitzendorf an der Schmida, Straning - Retzbach, Röschitz, Schrattenthal, Grafenberg, Zellerndorf Seefeld - Kadolz, Sigmundsherberg, Sitzendorf an der Schmida, Straning - Grafenberg, Zellerndorf, Ziersdorf Höhenstufen 438 m/ 226 m 468 m/ 191 m (max./min. m Höhe) * Quelle: Feinabgrenzung, Stand Mai 07 Die Europaschutzgebiete „Westliches Weinviertel” (FFH-Gebiet + Vogelschutzgebiet) sind Teil der Hauptregion Weinviertel. Das FFH-Gebiet besteht aus vielen kleinen Teilgebieten östlich des Manhartsberg-Zuges. Im Gegensatz dazu bildet das Vogelschutzgebiet ein weitgehend zusammenhängendes und deutlich größeres Gebiet, das sich bis in das Pulkautal im Norden ausdehnt. Das Gesamtgebiet (FFH-Gebiet + Vogelschutzgebiet) hat Anteil sowohl an der Böhmischen Masse mit ihren sauren, silikatischen Gesteinen, als auch an kalkhältigen Lössgebieten im östlichen Teil des Schutzgebiets. Das Gebiet ist -

The Reaction of Different Sorbus L. Species to Low Temperatures During Thaw in the Orel Region
Original Paper Journal of Forest Science, 65, 2019 (6): 218–225 https://doi.org/10.17221/8/2019-JFS The reaction of different Sorbus L. species to low temperatures during thaw in the Orel region Zoya Ozherelieva*, Olga Emelianova Russian Research Institute of Fruit Crop Breeding (VNIISPK), Orel, Russia Corresponding author: [email protected] *Citation: Ozherelieva Z., Emelianova O. (2019): The reaction of different Sorbus L. species to low temperatures during thaw in the Orel region. J. For. Sci., 65: 218–225. Abstract: Five Sorbus L. species of different ecological and geographical origin growing in the VNIISPK arboretum were studied. The Institute is located 368 km southwest of Moscow, on the Central Russian upland in the European part of Russia. The studies were carried out in 2014–2016. The reaction of different Sorbus L. species to a three-day thaw +2°C with a subsequent decrease in temperature to –25°C in February and –30°C in March was studied in or- der to identify adapted species to the climatic conditions of the Orel region for use in ornamental horticulture. As a result of the experiment, we recommend Sorbus aria (L.) Crantz, Sorbus aucuparia L. and Sorbus alnifolia (Siebold. et Zucc.) K. Koch. as adapted species for the Orel region to create sustainable landscape compositions. Keywords: artificial freezing; adaptation; frost hardiness Low temperature is a key environmental fac- es (Groffman et al. 2001; Schaberg et al. 2008; tor determining the evolution and distribution of Ozherelieva, Sedov 2017). Thus in nature, slight plants (Hawkins et al. 2014). Frost can damage freezing or death of trees is observed as a conse- plants through xylem embolism and the forma- quence of sharp temperature declines during thaws tion of extracellular ice, which causes cell dehydra- in February and March which cause trees to break tion and disruption of cell membranes (Zwiazek deep dormancy.