Dynamics of Pure Parasystole
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sick Sinus Syndrome in Children
Arch Dis Child: first published as 10.1136/adc.51.2.100 on 1 February 1976. Downloaded from Archives of Disease in Childhood, 1976, 51, 100. Sick sinus syndrome in children OLIVE SCOTT, FERGUS J. MACARTNEY, and PHILIP B. DEVERALL From the Department of Paediatric Cardiology,Killingbeck Hospital, Leeds Scott, O., Macartney, F. J., and Deverall, P. B. (1976). Archives of Disease in Childhood, 51, 100. Sick sinus syndrome in children. The ages of 6 male patients with the sick sinus syndrome ranged from 10-15 years when their symptoms began. At rest all had a heart rate of 60/min or less. Two had syncopal attacks which threatened life; 1 had only attacks of dizziness; the other 3 had no syncopal attacks but had recurrent attacks of supraventricular tachycardia ('brady-tachycardia syndrome') which were more resistant to drug therapy than is usual in childhood. They were not controlled or suppressed by digoxin when it was given. Substernal pain occurred in 2 patients who had syncope. In all patients the heart rate remained inappropriately slow after exercise and atropine. Cardiac pacemakers were used in the 2 patients with life-threatening syncope. Any patient who has dizziness or syncopal attacks and an inappropriately slow heart rate should have electrocardiograms recorded at rest and after excerise to record the heart rate and to look for abnormal P-waves. Dysfunction of the sinoatrial node has been sudden death in childhood than has been previously copyright. increasingly recognized over the past few years. recognized. An awareness of the condition may Most reported cases have been in adults and only a result in more cases being diagnosed. -

Basic Rhythm Recognition
Electrocardiographic Interpretation Basic Rhythm Recognition William Brady, MD Department of Emergency Medicine Cardiac Rhythms Anatomy of a Rhythm Strip A Review of the Electrical System Intrinsic Pacemakers Cells These cells have property known as “Automaticity”— means they can spontaneously depolarize. Sinus Node Primary pacemaker Fires at a rate of 60-100 bpm AV Junction Fires at a rate of 40-60 bpm Ventricular (Purkinje Fibers) Less than 40 bpm What’s Normal P Wave Atrial Depolarization PR Interval (Normal 0.12-0.20) Beginning of the P to onset of QRS QRS Ventricular Depolarization QRS Interval (Normal <0.10) Period (or length of time) it takes for the ventricles to depolarize The Key to Success… …A systematic approach! Rate Rhythm P Waves PR Interval P and QRS Correlation QRS Rate Pacemaker A rather ill patient……… Very apparent inferolateral STEMI……with less apparent complete heart block RATE . Fast vs Slow . QRS Width Narrow QRS Wide QRS Narrow QRS Wide QRS Tachycardia Tachycardia Bradycardia Bradycardia Regular Irregular Regular Irregular Sinus Brady Idioventricular A-Fib / Flutter Bradycardia w/ BBB Sinus Tach A-Fib VT PVT Junctional 2 AVB / II PSVT A-Flutter SVT aberrant A-Fib 1 AVB 3 AVB A-Flutter MAT 2 AVB / I or II PAT PAT 3 AVB ST PAC / PVC Stability Hypotension / hypoperfusion Altered mental status Chest pain – Coronary ischemic Dyspnea – Pulmonary edema Sinus Rhythm Sinus Rhythm P Wave PR Interval QRS Rate Rhythm Pacemaker Comment . Before . Constant, . Rate 60-100 . Regular . SA Node Upright in each QRS regular . Interval =/< leads I, II, . Look . Interval .12- .10 & III alike .20 Conduction Image reference: Cardionetics/ http://www.cardionetics.com/docs/healthcr/ecg/arrhy/0100_bd.htm Sinus Pause A delay of activation within the atria for a period between 1.7 and 3 seconds A palpitation is likely to be felt by the patient as the sinus beat following the pause may be a heavy beat. -

A Comparison of T-Wave Alternans, Signal Averaged Electrocardiography and Programmed Ventricular Stimulation for Arrhythmia Risk Stratification Michael R
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Journal of the American College of Cardiology Vol. 36, No. 7, 2000 © 2000 by the American College of Cardiology ISSN 0735-1097/00/$20.00 Published by Elsevier Science Inc. PII S0735-1097(00)01017-2 A Comparison of T-Wave Alternans, Signal Averaged Electrocardiography and Programmed Ventricular Stimulation for Arrhythmia Risk Stratification Michael R. Gold, MD, PHD, FACC,* Daniel M. Bloomfield, MD, FACC,† Kelley P. Anderson, MD, FACC,‡ Nabil E. El-Sherif, MD, FACC,§ David J. Wilber, MD, FACC, William J. Groh, MD, FACC,†† N. A. Mark Estes, III, MD, FACC,# Elizabeth S. Kaufman, MD, FACC,†† Mark L. Greenberg, MD, FACC,** David S. Rosenbaum, MD, FACC** Baltimore, Maryland; New York, and Brooklyn, New York; Pittsburgh, Pennsylvania; Chicago, Illinois; Indianapolis, Indiana; Boston, Massachusetts; Cleveland, Ohio; Lebanon, New Hampshire OBJECTIVES The goal of this study was to compare T-wave alternans (TWA), signal-averaged electro- cardiography (SAECG) and programmed ventricular stimulation (EPS) for arrhythmia risk stratification in patients undergoing electrophysiology study. BACKGROUND Accurate identification of patients at increased risk for sustained ventricular arrhythmias is critical to prevent sudden cardiac death. T-wave alternans is a heart rate dependent measure of repolarization that correlates with arrhythmia vulnerability in animal and human studies. Signal-averaged electrocardiography and EPS are more established tests used for risk stratification. METHODS This was a prospective, multicenter trial of 313 patients in sinus rhythm who were undergoing electrophysiologic study. T-wave alternans, assessed with bicycle ergometry, and SAECG were measured before EPS. -

Cardiology- the ABC's of the PQRST
Cardiology- the ABC’s of the PQRST Heather Carter LVT, VTS (Anesthesia & Analgesia) Introduction Understanding cardiology can provide the anesthetist with the ability to trouble shoot and treat various problems. Knowing when and how to treat a bradycardia versus a tachycardia can ensure a smooth anesthetic event. Origin of the Heartbeat http://i.pinimg.com/736x/9b/70/59/9b7059c20a452fc655a26e9e3e4b40ad.jpg Understanding the origin of the heartbeat can help the anesthetist know when and how to identify a potential problem. An ECG complex consists of a PQRST complex. The sinoatrial node (SA) is the pacemaker of the heart and produces the P wave. The QRS wave is produced by the atrioventricular node (AV). The P wave in an ECG complex indicates atrial depolarization. The QRS is responsible for ventricular depolarization and the T wave is ventricular repolarization. If a P wave is absent there is a lack of atrial depolarization. This is also known as atrial standstill. P waves that are combined in the QRS complexes are indicative of ventricular tachycardia or a junctional tachycardia. Present P waves that are without a QRS are indicative of an atrial depolarization that has not been conducted through the AV node. A QRS complex without a P wave demonstrate premature or escape beats. Essentials Arrhythmias can be identified with ease if several essentials are in place. ECG leads should be placed correctly: White- Right forelimb Black- Left forelimb Green- Right hind limb Red- Left hind limb Determining the paper speed (25mm/sec vs 50mm/sec) will also help the anesthetist determine if a bradycardia or tachycardia is present. -

T Wave Alternans After Sotalol: Evidence for Increased Sensitivity to Sotalol After Conversion from Atrial fibrillation to Sinus Rhythm
Heart 1998;80:303–306 303 CASE REPORT Heart: first published as 10.1136/hrt.80.3.303 on 1 September 1998. Downloaded from T wave alternans after sotalol: evidence for increased sensitivity to sotalol after conversion from atrial fibrillation to sinus rhythm H L Tan, A A M Wilde Abstract inhomogeneous distribution of repolari- A 64 year old woman with an 11 year sation controlling ion channels to induce Department of history of paroxysmal atrial fibrillation repolarisation abnormalities that may Cardiology, University of Amsterdam, presented to the emergency room because lead to torsade de pointes. Academic Medical of palpitations that had started two weeks (Heart 1998;80:303–306) Center, Amsterdam, previously. She had used sotalol 80 mg Netherlands once daily for three years without any epi- Keywords: T wave alternans; long QT syndrome; HLTan sodes of proarrhythmia or other adverse torsade de pointes; sotalol; atrial fibrillation Department of eVects. However, she developed pro- Cardiology, Heart nounced T wave alternans with giant Lung Institute, Utrecht inverted T waves and excessive QT pro- The ECG T wave reflects cardiac repolarisa- University Hospital, longation following sotalol administration tion. Its broad inscription results from the tem- Utrecht, Netherlands one day after conversion from atrial A A M Wilde poral inhomogeneity with which repolarisation fibrillation to sinus rhythm. This case proceeds through the heart. T wave changes, demonstrates bizarre T wave changes, T Correspondence to: including T wave inversion and T wave altern- Dr H L Tan, Department of wave alternans, and extreme QT prolon- ans, have been subdivided into primary and Cardiology, Academic gation following sotalol administration Medical Center, secondary forms. -

Basic Cardiac Rhythms – Identification and Response Module 1 ANATOMY, PHYSIOLOGY, & ELECTRICAL CONDUCTION Objectives
Basic Cardiac Rhythms – Identification and Response Module 1 ANATOMY, PHYSIOLOGY, & ELECTRICAL CONDUCTION Objectives ▪ Describe the normal cardiac anatomy and physiology and normal electrical conduction through the heart. ▪ Identify and relate waveforms to the cardiac cycle. Cardiac Anatomy ▪ 2 upper chambers ▪ Right and left atria ▪ 2 lower chambers ▪ Right and left ventricle ▪ 2 Atrioventricular valves (Mitral & Tricuspid) ▪ Open with ventricular diastole ▪ Close with ventricular systole ▪ 2 Semilunar Valves (Aortic & Pulmonic) ▪ Open with ventricular systole ▪ Open with ventricular diastole The Cardiovascular System ▪ Pulmonary Circulation ▪ Unoxygenated – right side of the heart ▪ Systemic Circulation ▪ Oxygenated – left side of the heart Anatomy Coronary Arteries How The Heart Works Anatomy Coronary Arteries ▪ 2 major vessels of the coronary circulation ▪ Left main coronary artery ▪ Left anterior descending and circumflex branches ▪ Right main coronary artery ▪ The left and right coronary arteries originate at the base of the aorta from openings called the coronary ostia behind the aortic valve leaflets. Physiology Blood Flow Unoxygenated blood flows from inferior and superior vena cava Right Atrium Tricuspid Valve Right Ventricle Pulmonic Valve Lungs Through Pulmonary system Physiology Blood Flow Oxygenated blood flows from the pulmonary veins Left Atrium Mitral Valve Left Ventricle Aortic Valve Systemic Circulation ▪ Blood Flow Through The Heart ▪ Cardiology Rap Physiology ▪ Cardiac cycle ▪ Represents the actual time sequence between -

Revisiting Electrocardiographic Wolff-Parkinson-White Pattern
The Journal of Medical Research 2020; 6(4): 114-116 Review Article Revisiting Electrocardiographic Wolff-Parkinson-White Pattern JMR 2020; 6(4): 114-116 1 2 July- August Pradnya Brijmohan Bhattad , Vinay Jain ISSN: 2395-7565 1 Resident, Department of Internal Medicine, East Tennessee State University, Tennessee (TN), USA © 2020, All rights reserved 2 Attending Radiologist, Department of Radiology, James H. Quillen Veterans Affairs Medical Center, Mountain www.medicinearticle.com Home, Tennessee (TN), USA Received: 14-06-2020 Accepted: 18-07-2020 Abstract Electrocardiographic features of a short PR interval, a delta wave, and a wide QRS complex constitutes a Wolff- Parkinson-White (WPW) pattern. Asymptomatic electrocardiographic findings are defined as a WPW pattern. Symptomatic patients with these electrocardiographic features have WPW syndrome. WPW syndrome may predispose to arrhythmias such as paroxysmal atrial tachycardia, atrial fibrillation, ventricular tachycardia. Patient’s with WPW syndrome at risk for sudden cardiac death. It is important to recognize the common electrocardiographic characteristics of WPW pattern. The advent of electrophysiological studies (EPS) and radiofrequency ablation has revolutionized the management of WPW syndrome. Keywords: Wolff-Parkinson-White Syndrome, Wolff-Parkinson-White Pattern, Pre-excitation, Accessory pathway. INTRODUCTION Wolff, Parkinson, and White described a patient series in the year 1930 who suffered from paroxysms of tachycardia with classic electrocardiographic findings which has been described as WPW pattern [1]. It is a congenital abnormality in the cardiac conduction system. WPW pattern is a ventricular pre-excitation entity wherein an accessory bypass tract known as the bundle of Kent serves as the connection between the atrial to the ventricular myocardium bypassing the atrioventricular (AV) node [2,3]. -

Management of Wolff-Parkinson-White
Management of Wolff-Parkinson-White Tachyarrhythmia Presenting as Syncope with Seizure- like Activity * * Samuel Kaplan, BS and Lindsey Spiegelman, MD *University of California, Irvine, Department of Emergency Medicine, Orange, CA Correspondence should be addressed to Samuel Kaplan at [email protected] Submitted: August 15, 2017; Accepted: September 13, 2017; Electronically Published: October 15, 2017; https://doi.org/10.21980/J8534P Copyright: © 2017 Kaplan, et al. This is an open access article distributed in accordance with the terms of the Creative Commons Attribution (CC BY 4.0) License. See: http://creativecommons.org/licenses/by/4.0/ Empty Line Calibri Size 12 Empty Line Calibri Size 12 ABSTRACT: Audience: Emergency medicine residents and medical students. Introduction: An estimated 3% of the United States population suffers from recurrent convulsive episodes that are most often attributed to primary epileptic seizures.1 However, recent studies have estimated about 20%-30% of such episodes are associated with occult cardiac etiology,2 which carry one-year mortality rates of up to 30%.3 Cardiogenic cerebral hypoxia has been associated with a wide variety of neurologic disturbances, including dizzy spells, headache, syncope, focal motor deficit, generalized tonic-clonic seizure, confusion, dementia, and psychosis.4 Convulsive activity has tentatively been ascribed to the ensuing activation of the medullary reticular formation.5,6 This scenario is based on a patient that presented to University of California Irvine Medical Center Emergency -

Sinus Bradycardia
British Heart Journal, I97I, 33, 742-749. Br Heart J: first published as 10.1136/hrt.33.5.742 on 1 September 1971. Downloaded from Sinus bradycardia Dennis Eraut and David B. Shaw From the Cardiac Department, Royal Devon and Exeter Hospital, Exeter, Devon This paper presents thefeatures of 46 patients with unexplained bradycardia. Patients were ad- mitted to the study if their resting atrial rate was below 56 a minute on two consecutive occasions. Previous electrocardiograms and the response to exercise, atropine, and isoprenaline were studied. The ages of thepatients variedfrom I3 to 88years. Only 8 had a past history ofcardiovascular disease other than bradycardia, but 36 hJd syncopal or dizzy attacks. Of the 46 patients, 35 had another arrhythmia in addition to bradycardia; at some stage, i6 had sinus arrest, i.5 hadjunc- tional rhythm, 12 had fast atrial arrhythmia, I6 had frequent extrasystoles, and 6 had atrio- ventricular block. None had the classical features of sinoatrial block. Arrhythmias were often produced by exercise, atropine, or isoprenaline. Drug treatment was rarely satisfactory, but only i patient needed a permanent pacemaker. It is suggested that the majority of the patients were suffering from a pathological form of sinus bradycardia. The aetiology remains unproven, but the most likely explanation is a loss of the inherent rhythmicity of the sinoatrial node due to a primary degenerative disease. The descriptive title of 'the lazy sinus syndrome' is suggested. copyright. Bradycardia with a slow atrial rate is usually attempt to define the clinical syndrome of regarded as an innocent condition common in bradycardia with a pathologically slow atrial certain types of well-trained athlete, but occa- rate and to clarify the nature of the arrhyth- sionally it may occur in patients with symp- mia. -
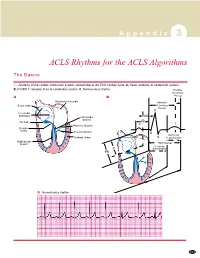
ACLS Rhythms for the ACLS Algorithms
A p p e n d i x 3 ACLS Rhythms for the ACLS Algorithms The Basics 1. Anatomy of the cardiac conduction system: relationship to the ECG cardiac cycle. A, Heart: anatomy of conduction system. B, P-QRS-T complex: lines to conduction system. C, Normal sinus rhythm. Relative Refractory A B Period Bachmann’s bundle Absolute Sinus node Refractory Period R Internodal pathways Left bundle AVN branch AV node PR T Posterior division P Bundle of His Anterior division Q Ventricular Purkinje fibers S Repolarization Right bundle branch QT Interval Ventricular P Depolarization PR C Normal sinus rhythm 253 A p p e n d i x 3 The Cardiac Arrest Rhythms 2. Ventricular Fibrillation/Pulseless Ventricular Tachycardia Pathophysiology ■ Ventricles consist of areas of normal myocardium alternating with areas of ischemic, injured, or infarcted myocardium, leading to chaotic pattern of ventricular depolarization Defining Criteria per ECG ■ Rate/QRS complex: unable to determine; no recognizable P, QRS, or T waves ■ Rhythm: indeterminate; pattern of sharp up (peak) and down (trough) deflections ■ Amplitude: measured from peak-to-trough; often used subjectively to describe VF as fine (peak-to- trough 2 to <5 mm), medium-moderate (5 to <10 mm), coarse (10 to <15 mm), very coarse (>15 mm) Clinical Manifestations ■ Pulse disappears with onset of VF ■ Collapse, unconsciousness ■ Agonal breaths ➔ apnea in <5 min ■ Onset of reversible death Common Etiologies ■ Acute coronary syndromes leading to ischemic areas of myocardium ■ Stable-to-unstable VT, untreated ■ PVCs with -

Microvolt T-Wave Alternans (MTWA) Warns of Impending Cardiac Arrest
and reported shortness of breath but denied chest Microvolt Twave Alternans (MTWA) pain. Approximately two and half minutes into the Warns of Impending Cardiac Arrest recovery phase, the patient developed polymorphic Parag V. Patel, DO, FACC, Advocate Lutheran General ventricular tachycardia which quickly deteriorated Hospital, Park Ridge, Illinois into ventricular fibrillation (VF) (Figure 1). An AED was applied within three minutes and the Introduction patient was successfully defibrillated on the second T‐wave alternans (TWA) refers to an alternating attempt. pattern in the T‐wave of the electrocardiogram (ECG) and is recognized as a marker of electrical Upon review of the test, ST segment changes instability. Visible or macroscopic alternans was suggestive of ischemia were evident, but there first reported in the context of proarrhythmic were no urgent symptoms or overt indications on conditions including Prinzmetal’s angina and long the ECG that signaled the impending arrhythmia QT syndrome, and was often found to immediately (e.g., ventricular ectopy or visible repolarization precede the onset of ventricular tachycardia or alternans). From the MTWA report, however, it is fibrillation [1‐3]. More recently, techniques have clear that non‐visible microvolt‐level T‐wave been developed to measure microvolt‐level T‐wave alternans was present during the exercise portion alternans (MTWA) not visible to the human eye [4]. of the test and immediately preceded the onset of The presence of MTWA has been linked with VF (Figure 2). increased risk of sudden cardiac arrest (SCA) in several patient populations [5‐8]. This study MTWA detected during stress testing presents the case of a patient who experienced VF may indicate the presence of active during the recovery phase of an MTWA stress test. -

European Heart Rhythm Association (EHRA) Consensus Document on the Management of Supraventricular Arrhythmias, Endorsed by Heart
Europace (2017) 19, 465–511 EHRA POSITION PAPER doi:10.1093/europace/euw301 European Heart Rhythm Association (EHRA) consensus document on the management of supraventricular arrhythmias, endorsed by Heart Rhythm Society (HRS), Asia-Pacific Heart Downloaded from https://academic.oup.com/europace/article/19/3/465/2631183 by guest on 18 August 2020 Rhythm Society (APHRS), and Sociedad Latinoamericana de Estimulacio´nCardiacay Electrofisiologia (SOLAECE) Demosthenes G. Katritsis, (Chair)1, Giuseppe Boriani2,FranciscoG.Cosio3, Gerhard Hindricks4,PierreJaı¨s5,MarkE.Josephson6, Roberto Keegan7, Young-Hoon Kim8, Bradley P. Knight9,Karl-HeinzKuck10,DeirdreA.Lane10,11, GregoryY.H.Lip11, Helena Malmborg12, Hakan Oral13, Carlo Pappone14, Sakis Themistoclakis15,KathrynA.Wood16, and Carina Blomstro¨m-Lundqvist, (Co-Chair)12 REVIEWERS: Bulent Gorenek, (review coordinator)17, Nikolaos Dagres4, Gheorge-Andrei Dan18, Marc A Vos19, Gulmira Kudaiberdieva20, Harry Crijns21, Kurt Roberts-Thomson22, Yenn-Jiang Lin23, Diego Vanegas24, Walter Reyes Caorsi25, Edmond Cronin26, and Jack Rickard27 1Athens Euroclinic, Athens, Greece; and Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA, USA; 2Cardiology Department, Modena University Hospital, University of Modena and Reggio Emilia, Modena, Italy; 3Hospital Universitario De Getafe, Madrid, Spain; 4University of Leipzig, Heartcenter, Leipzig, Germany; 5University of Bordeaux, CHU Bordeaux, LIRYC, France; 6Berth Israel Deaconess Medical Center, Boston, MA, USA; 7Hospital Privado del Sur y Hospital