J. APIC. SCI. Vol. 63 No. 2 2019 DOI: 10.2478/JAS-2019-0022
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ETHIOPIA - National Hot Spot Map 31 May 2010
ETHIOPIA - National Hot Spot Map 31 May 2010 R Legend Eritrea E Tigray R egion !ª D 450 ho uses burned do wn d ue to th e re ce nt International Boundary !ª !ª Ahferom Sudan Tahtay Erob fire incid ent in Keft a hum era woreda. I nhabitan ts Laelay Ahferom !ª Regional Boundary > Mereb Leke " !ª S are repo rted to be lef t out o f sh elter; UNI CEF !ª Adiyabo Adiyabo Gulomekeda W W W 7 Dalul E !Ò Laelay togethe r w ith the regiona l g ove rnm ent is Zonal Boundary North Western A Kafta Humera Maychew Eastern !ª sup portin g the victim s with provision o f wate r Measle Cas es Woreda Boundary Central and oth er imm ediate n eeds Measles co ntinues to b e re ported > Western Berahle with new four cases in Arada Zone 2 Lakes WBN BN Tsel emt !A !ª A! Sub-city,Ad dis Ababa ; and one Addi Arekay> W b Afa r Region N b Afdera Military Operation BeyedaB Ab Ala ! case in Ahfe rom woreda, Tig ray > > bb The re a re d isplaced pe ople from fo ur A Debark > > b o N W b B N Abergele Erebtoi B N W Southern keb eles of Mille and also five kebeles B N Janam ora Moegale Bidu Dabat Wag HiomraW B of Da llol woreda s (400 0 persons) a ff ected Hot Spot Areas AWD C ases N N N > N > B B W Sahl a B W > B N W Raya A zebo due to flo oding from Awash rive r an d ru n Since t he beg in nin g of th e year, Wegera B N No Data/No Humanitarian Concern > Ziquala Sekota B a total of 967 cases of AWD w ith East bb BN > Teru > off fro m Tigray highlands, respective ly. -

Ethiopia-Kenya
THE FEDERAL DEMOCRATIC REPUBLIC OF KENYA REPUBLIC OF ETHIOPIA ETHIOPIAN ELECTRIC POWER KENYA ELECTRICITY TRANSMISSION COMPANY LIMITED CORPORATION (EEPCo) ETHIOPIA-KENYA POWER SYSTEMS INTERCONNECTION PROJECT REVISION OF ENVIRONMENTAL AND SOCIAL IMPACT ASSESSMENT AND RESETTLEMENT ACTION PLAN STUDIES RESETTLEMENT ACTION PLAN (RAP) FINAL REPORT PART 1: ETHIOPIA JANUARY 2012 Tropics Consulting Engineers Plc Gamma Systems Ltd P.O.Box 351 Addis Ababa, Ethiopia P O Box 1033 – 00606 TEL 251-11-618 54 66 Fax 251-11-618 38 61 NAIROBI, Kenya Tel: +254 20 44 51 528 e-mail: [email protected] Fax: +254 20 44 51 529 web-site www.tropicsconsultingengineers.com Ethiopian Electric Power Corporation (EEPCo) Ethiopia-Kenya Power Systems Interconnection Project Kenya Electricity Transmission Company Limited Resettlement Action Plan (RAP) Final Report General Table of Contents Pages E. Executive Summary ..................................................................................... E-0 1 Introduction .................................................................................................. 1-1 2. Description of the project, project area and area of influence ...................... 2-1 3 Potential Impacts ......................................................................................... 3-1 4. Organizational Responsibility ....................................................................... 4-1 5 Community Participation .............................................................................. 5-1 6 Integration with host communities -

Challenges and Contributions of Crop Production in Agro-Pastoral Systems of Borana Plateau, Ethiopia Abera Tilahun1*, Beyene Teklu2 and Dana Hoag3
Tilahun et al. Pastoralism: Research, Policy and Practice (2017) 7:11 Pastoralism: Research, Policy DOI 10.1186/s13570-017-0088-y and Practice ERRATUM Open Access Erratum to: Challenges and contributions of crop production in agro-pastoral systems of Borana Plateau, Ethiopia Abera Tilahun1*, Beyene Teklu2 and Dana Hoag3 Erratum southern Ethiopia, unpublished; Oba, G. 1998: Assess- ment of indigenous range management knowledge of Abstract the Booran pastoralists of southern Ethiopia). At the This erratum expands on the original article by Tilahun policy level, this fact is also generally accepted and ac- et al. (2017) published on 16 February 2017, and seeks knowledged as one of the strategies for income diversifi- to correct any oversights in the original article. Only the cation of the pastoralists and the support for cropping 'edited' paragraphs are available in this Erratum, the full is likely to continued (Mohammed, M., 2004: A com- original article can be accessed via: DOI 10.1186/ parative study of pastoralist parliamentary groups: s13570-016-0074-9. Case study on the pastoral affairs standing committee of Ethiopia, unpublished). Introduction In the original paper 2 objectives have been stated, but In the introduction the relation between crop produc- these should have been the following 3: (i) to identify tion and pastoralism is stated, the detailed introduction major determinants of crop production in pastoralist is as followed: households, (ii) to assess the contribution of crop produc- Followed these changes, pastoralists seek additional tion to the household economy of agro-pastoral commu- ways to earn a living as they watch historical pastoralist nities, and. -

Oromia Region Administrative Map(As of 27 March 2013)
ETHIOPIA: Oromia Region Administrative Map (as of 27 March 2013) Amhara Gundo Meskel ! Amuru Dera Kelo ! Agemsa BENISHANGUL ! Jangir Ibantu ! ! Filikilik Hidabu GUMUZ Kiremu ! ! Wara AMHARA Haro ! Obera Jarte Gosha Dire ! ! Abote ! Tsiyon Jars!o ! Ejere Limu Ayana ! Kiremu Alibo ! Jardega Hose Tulu Miki Haro ! ! Kokofe Ababo Mana Mendi ! Gebre ! Gida ! Guracha ! ! Degem AFAR ! Gelila SomHbo oro Abay ! ! Sibu Kiltu Kewo Kere ! Biriti Degem DIRE DAWA Ayana ! ! Fiche Benguwa Chomen Dobi Abuna Ali ! K! ara ! Kuyu Debre Tsige ! Toba Guduru Dedu ! Doro ! ! Achane G/Be!ret Minare Debre ! Mendida Shambu Daleti ! Libanos Weberi Abe Chulute! Jemo ! Abichuna Kombolcha West Limu Hor!o ! Meta Yaya Gota Dongoro Kombolcha Ginde Kachisi Lefo ! Muke Turi Melka Chinaksen ! Gne'a ! N!ejo Fincha!-a Kembolcha R!obi ! Adda Gulele Rafu Jarso ! ! ! Wuchale ! Nopa ! Beret Mekoda Muger ! ! Wellega Nejo ! Goro Kulubi ! ! Funyan Debeka Boji Shikute Berga Jida ! Kombolcha Kober Guto Guduru ! !Duber Water Kersa Haro Jarso ! ! Debra ! ! Bira Gudetu ! Bila Seyo Chobi Kembibit Gutu Che!lenko ! ! Welenkombi Gorfo ! ! Begi Jarso Dirmeji Gida Bila Jimma ! Ketket Mulo ! Kersa Maya Bila Gola ! ! ! Sheno ! Kobo Alem Kondole ! ! Bicho ! Deder Gursum Muklemi Hena Sibu ! Chancho Wenoda ! Mieso Doba Kurfa Maya Beg!i Deboko ! Rare Mida ! Goja Shino Inchini Sululta Aleltu Babile Jimma Mulo ! Meta Guliso Golo Sire Hunde! Deder Chele ! Tobi Lalo ! Mekenejo Bitile ! Kegn Aleltu ! Tulo ! Harawacha ! ! ! ! Rob G! obu Genete ! Ifata Jeldu Lafto Girawa ! Gawo Inango ! Sendafa Mieso Hirna -

190327 Oromia Region Agric S
ETHIOPIA: AGRICULTURE SECTOR HRP OROMIA REGION MONTHLY DASHBOARD - March 2019 The devastating impact on agriculture following consecutive years of drought in Ethiopia is undisputed. While forecasts for 2019 indicate a probability of normal to above normal rain in most parts of Ethiopia, in east, south and southeastern regions, the upcoming rainy season (March to June) is forecast- KEY FIGURES ed to be average or below average. In areas where normal to above normal rains are expected, recovery will not be spontaneous, as previous OVERVIEW HOUSEHOLDS REACHED drought-affected households are likely to require sustained humanitarian assistance as a result of exhausted coping mechanisms. HOUSEHOLDS IN NEED Humanitarian assistance for IDPs and IDP returnees is largely dependent on IDPs’ access to land and the livelihood assets they have been able to 1.15 million maintain during displacement. Emergency feed and animal health interventions are needed to reduce the burden on the resources of the host 0.0m 0% communities and prevent the spread of diseases, especially for animals displaced across regional borders. Where appropriate, land will be availed and crop seeds, farming tools, and training will be provided to support IDP and returning households to improve their food security and reduce the burden HOUSEHOLDS TARGETED on host Communities. 658,428 IDP HOUSEHOLDS TARGETED N_Shewa 0m 0% Horo 64,195 Guduru Chinaksen Guto W_Wellega Gida Meta Sasiga Finfine Doba Gursum AGRICULTURE LIVESTOCK E_Wellega Mieso Crop Seeds &Tools W_Shewa Special Girawa -

Damage and Loss Assessment EXECUTIVE SUMMARY ©UNHCR/Adelina Gomez
©UNHCR/Anna Helge Damage and Loss Assessment EXECUTIVE SUMMARY ©UNHCR/Adelina Gomez TABLE OF CONTENT EXECUTIVE SUMMARY 3 DAMAGE AND LOSS IN AGRICULTURAL PRODUCTION SECTORS 4 DAMAGE ON LIVESTOCK PRODUCTION SYSTEM 5 DAMAGE AND LOSS ASSESMENT ON SOCIAL AND INFRAESTRUCTURAL 6 SECTORS • Private houses 6 • Public Infrastructure 7 • Recommendations 8 LEGAL ASSISTANCE NEEDS 10 • Access to justice 10 • Access to Land holding certificate 11 • Customary/Traditional justice system 12 • Recommendations 12 CONFLICT, PEACE BUILDING AND SOCIAL COHESION 13 POSSIBLE APPROACHES FOR CONFLICT TRANSFORMATION AND PEACE- 14 BUILDING IN THE CONTEXT OF GUJI AND GEDEO ETHNIC GROUPS • Building Trust 14 • Developing social cohesion 15 • Supporting local buy-in 15 • Recommendations 16 APPENDIX 1 18 - 2 - EXECUTIVE SUMMARY Damage and Loss Assessment in West Guji Bule Hora University initiated a Damage and Loss Assessment (DaLA) in West Guji zone in collaboration with the United Nations High Commissioner for Refugees (UNHCR) and the West Guji Zonal Disaster Risk Management Office to assess loss and damages incurred during the intercommunal conflict between the Gedeo and Guji people in 2018, which displaced hundreds of thousands. The assessment also analyzed legal assistance needs, as well as the potential for peace-building and social cohesion. A total of 78,257HHs (508,671 individuals) with an average family size of 6.5/HH were severely exposed to multiple socio-economic crises due to the conflict. Following the Government’s initiated return process based on the alleged improved safety and security situation (assessed by the Government) in May 2019, according to the Government, the majority of the IDPs were returned to their places of habitual residence 1. -

Resettlement and Local Livelihoods in Nechsar National Park, Southern Ethiopia
Resettlement and Local Livelihoods in Nechsar National Park, Southern Ethiopia Abiyot Negera Biressu Thesis Submitted for the Degree: Master of Philosophy in Indigenous Studies Faculty of Social Science, University of Tromsø Norway, Spring 2009 Resettlement and Local Livelihoods in Nechsar National Park, Southern Ethiopia By: Abiyot Negera Biressu Thesis Submitted for the Degree: Master of Philosophy in Indigenous Studies Faculty of Social Sciences, University of Tromsø Norway Tromso, Spring 2009 Acknowledgement I would like to thank people and institutions that provided me with the necessary support for my education in Tromsø and during the production of this thesis. I am very much thankful to the Norwegian State Educational Loan Fund (Lånnekassen) for financing my education here at the University of Tromsø. My gratitude also goes to Center for Sámi Studies for financing my fieldwork. I would like to say, thank you, to my supervisor Ivar Bjørklund (Associate Professor) for his comments during the writing of this thesis. My friends (Ashenafi, Eba and Tariku), whose encouraging words are always a click away from me, also deserve special thanks. I am also indebted to my informants and Nechsar National Park Administration for their cooperation during the field work. i Table of Contents Acknowledgement…………………………………………………………………………………………...i Acronyms……………………………………………………………………………………………………v List of Maps………………………………………………………………...………………………………vi Abstract…………………………………………………………………………………………………….vii Chapter One: Introduction…………………………………………………………………………...........1 1.1. Introduction to the Place and People of Study Area…………………………………………………….1 1.2. Research Frame………………………………………………………………………………………….2 1.3. Objective and Significance of the Study………………………………………………………………...5 1.4. Methodology…………………………………………………………………………………………….5 1.4.1. From Park Management to Guji Community…………………………………………………………5 1.4.2. Oral Interview…………………………………………………………………………………………7 1.4.3. -
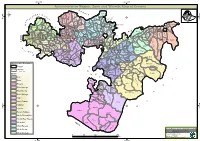
Administrative Region, Zone and Woreda Map of Oromia a M Tigray a Afar M H U Amhara a Uz N M
35°0'0"E 40°0'0"E Administrative Region, Zone and Woreda Map of Oromia A m Tigray A Afar m h u Amhara a uz N m Dera u N u u G " / m r B u l t Dire Dawa " r a e 0 g G n Hareri 0 ' r u u Addis Ababa ' n i H a 0 Gambela m s Somali 0 ° b a K Oromia Ü a I ° o A Hidabu 0 u Wara o r a n SNNPR 0 h a b s o a 1 u r Abote r z 1 d Jarte a Jarso a b s a b i m J i i L i b K Jardega e r L S u G i g n o G A a e m e r b r a u / K e t m uyu D b e n i u l u o Abay B M G i Ginde e a r n L e o e D l o Chomen e M K Beret a a Abe r s Chinaksen B H e t h Yaya Abichuna Gne'a r a c Nejo Dongoro t u Kombolcha a o Gulele R W Gudetu Kondole b Jimma Genete ru J u Adda a a Boji Dirmeji a d o Jida Goro Gutu i Jarso t Gu J o Kembibit b a g B d e Berga l Kersa Bila Seyo e i l t S d D e a i l u u r b Gursum G i e M Haro Maya B b u B o Boji Chekorsa a l d Lalo Asabi g Jimma Rare Mida M Aleltu a D G e e i o u e u Kurfa Chele t r i r Mieso m s Kegn r Gobu Seyo Ifata A f o F a S Ayira Guliso e Tulo b u S e G j a e i S n Gawo Kebe h i a r a Bako F o d G a l e i r y E l i Ambo i Chiro Zuria r Wayu e e e i l d Gaji Tibe d lm a a s Diga e Toke n Jimma Horo Zuria s e Dale Wabera n a w Tuka B Haru h e N Gimbichu t Kutaye e Yubdo W B Chwaka C a Goba Koricha a Leka a Gidami Boneya Boshe D M A Dale Sadi l Gemechis J I e Sayo Nole Dulecha lu k Nole Kaba i Tikur Alem o l D Lalo Kile Wama Hagalo o b r Yama Logi Welel Akaki a a a Enchini i Dawo ' b Meko n Gena e U Anchar a Midega Tola h a G Dabo a t t M Babile o Jimma Nunu c W e H l d m i K S i s a Kersana o f Hana Arjo D n Becho A o t -

Ethiopia: 3W - Health Cluster Ongoing Activities Map (December 2016)
Ethiopia: 3W - Health Cluster Ongoing Activities map (December 2016) ERITREA 8 Total Number of Partners Ahferom CCM CCM GOAL GOAL Erob CCM Adwa GOAL Red Sea GOAL Werei CCM Leke GOAL Koneba GOAL Hawzen GOAL CCM SUDAN TIGRAY GOAL Ab Ala GOAL AMHARA Megale Gulf of GOAL Aden DCA IMC Kobo AFAR Lay DCA Meket DCA Gayint IMC IMC Tach Gayint DCA Guba Lafto GOAL BENESHANGUL Dera IMC Worebabu Simada GOAL GOAL GOAL GOAL GUMU IMC Thehulederie Sirba DJIBOUTI Abay Telalak Afambo GOAL GOAL IRC Tenta GOAL Sayint GOAL GOAL IRC GOAL GOAL Were Ilu Ayisha IRC IRC GOAL Dewa Sherkole Legehida Harewa Kurmuk GOAL IMC Menge Kelela Artuma IRC Yaso Fursi IMC Erer IRC IRC IRC Jille Menz IMC Timuga Dembel Wara Afdem Bilidigilu IRC Mama Assosa IRC Jarso IMC Tarema IMC Midir Ber IRC IRC Agalometi Gerar IMC Jarso Kamashi IMC Bambasi GOAL DIRE Chinaksen IMC IMC IRC IMC DAWA IMC Bio Jiganifado Ankober Meta IRC GOAL IRC IMC IMC Aleltu Deder HARERI GOAL GOAL Gursum IRC IRC IRC GOAL Midega SOMALIA IRC IMC Goba SOUTH SUDAN Tola ACF Koricha Anfilo IMC Gashamo Anchar GOAL Daro Lebu Boke Golo Oda IRC Wantawo GOAL Meyu IMC IRC IRC IRC GOAL GOAL IMC Aware SCI IMC Fik IRC IRC Kokir Sire Jikawo IRC Gedbano Adami IMC GOAL Tulu Jido Degehabur GOAL SCI GOAL Sude Akobo Selti Kombolcha IRC IRC Lanfero Hamero Gunagado Mena Dalocha IMC GAMBELA GOAL Arsi IMC Shekosh GOAL Gololcha GOAL Negele Bale IMC Soro GOAL IMC IRC GOAL IMC Agarfa IRC Tembaro IRC IRC GOAL SCI GOAL GOAL IMC IMC Ginir CCM GOAL GOAL IRC IMC IMC GOAL GOAL IRC GOAL Sinana IMC IRC IRC Dinsho GOAL Goba IRC IMC GOAL IRC GOAL IRC Adaba CCM GOAL Berbere IMC Humbo GOAL SOMALI IMC Hulla IRC GOAL CCM GOAL GOAL GOAL PIN IRC Zala IMC IRC IRC Abaya PIN IRC Wenago Ubadebretsehay Mirab Gelana Abaya IRC GOAL GOAL SCI IRC IRC SCI Amaro OROMIA SNNPR IRC SCI CCM Bonke GOAL IRC Meda CCM SCI Welabu Legend SCI Konso IMC SCI International boundary Filtu Hudet INDIAN Agencies' locOaCtiEoAnNs and Regional boundary SCI Arero Dolobay Dolo Odo area of interventions are IMC No. -

Periodic Monitoring Report Working 2016 Humanitarian Requirements Document – Ethiopia Group
DRMTechnical Periodic Monitoring Report Working 2016 Humanitarian Requirements Document – Ethiopia Group Covering 1 Jan to 31 Dec 2016 Prepared by Clusters and NDRMC Introduction The El Niño global climactic event significantly affected the 2015 meher/summer rains on the heels of failed belg/ spring rains in 2015, driving food insecurity, malnutrition and serious water shortages in many parts of the country. The Government and humanitarian partners issued a joint 2016 Humanitarian Requirements Document (HRD) in December 2015 requesting US$1.4 billion to assist 10.2 million people with food, health and nutrition, water, agriculture, shelter and non-food items, protection and emergency education responses. Following the delay and erratic performance of the belg/spring rains in 2016, a Prioritization Statement was issued in May 2016 with updated humanitarian requirements in nutrition (MAM), agriculture, shelter and non-food items and education.The Mid-Year Review of the HRD identified 9.7 million beneficiaries and updated the funding requirements to $1.2 billion. The 2016 HRD is 69 per cent funded, with contributions of $1.08 billion from international donors and the Government of Ethiopia (including carry-over resources from 2015). Under the leadership of the Government of Ethiopia delivery of life-saving and life- sustaining humanitarian assistance continues across the sectors. However, effective humanitarian response was challenged by shortage of resources, limited logistical capacities and associated delays, and weak real-time information management. This Periodic Monitoring Report (PMR) provides a summary of the cluster financial inputs against outputs and achievements against cluster objectives using secured funding since the launch of the 2016 HRD. -

Ethiopian Coffee Buying Guide
Fintrac Inc. www.fintrac.com [email protected] US Virgin Islands 3077 Kronprindsens Gade 72 St. Thomas, USVI 00802 Tel: (340) 776-7600 Fax: (340) 776-7601 Washington, D.C. 1436 U Street NW, Suite 303 Washington, D.C. 20009 USA Tel: (202) 462-8475 Fax: (202) 462-8478 USAID’s Agribusiness and Trade Expansion Program 4th Floor, Ki-Ab Building Alexander Pushkin Street Kebele 23 Addis Ababa Ethiopia Tel: + 251-(0)11-372-0060/61/62/63/64 Fax: + 251-(0)11-372-0102 () parenthesis indicate use within Ethiopia only Author Willem J. Boot Photography All photos by Willem J. Boot March 2011 This publication was produced for review by the United States Agency for International Development. It was prepared by Willem J. Boot for Fintrac Inc., the U.S. contractor implementing USAID/Ethiopia’s Agribusiness and Trade Expansion Program. March 2011 The author’s views expressed in this publication do not necessarily reflect the views of the United States Agency for International Development or the United States Government. CONTENT INTRODUCTION ....................................................................................................................... 1 PART ONE: THE MOTHERLAND OF COFFEE ...................................................................... 2 Culture and Geography ................................................................................................ 2 Coffee Characteristics ................................................................................................. 3 Processing: Sun-dried Natural Coffees vs. Washed Coffees -

Urgent Need for Integrated Emergency Response in 141 Outbreak and Undernutrition Affected Woredas of Ethiopia in 2020
JOINT INTER-CLUSTER ADVOCACY PAPER URGENT NEED FOR INTEGRATED EMERGENCY RESPONSE IN 141 OUTBREAK AND UNDERNUTRITION AFFECTED WOREDAS OF ETHIOPIA IN 2020 Ethiopian population is currently facing new crises while past crises are not yet resolved. The recurrent food & undernutrition crises and increased frequency of measles and cholera outbreaks in the course of 2019 are now further exacerbated by desert locust infestations and upcoming COVID-19 pandemic. The complexification of the emergency needs requires to bring together our integrated efforts and expertise to address them. Ethiopia Health Cluster, WASH Cluster and Emergency Nutrition Coordination Unit established Health, WASH and Nutrition Technical Woking Group (HWN TWG) to further boost joint response in 141 woreda and IDP camps of Ethiopia by implementing minimum multi-sectoral response package. OUR ASKS • The HWN TWG asks the EHCT to support our initiative by providing more visibility and attention to this integrated initiative and request an opportunity to provide periodic updates at EHCT meetings. • The HWN TWG asks the Government of Ethiopia, especially NDRMC, MOH, EPHI and MOWIE, to improve the joint Health, WASH and Nutrition collaboration, to foster joint system strengthening and data sharing with humanitarian actors. • The HWN TWG asks Donors, including EHF and all the Rapid Response Mechanisms, to prioritise multisectoral funding in priority woredas and consider the multi-sectoral approach as an example of good practice. • The HWN TWG asks implementing partners to include the pilot of the Health WASH and Nutrition minimum package in their future proposals and project design. Partners are encouraged to work as a consortium to jointly implement the full package.