Review Questions DNA Replication
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Copyright by Young-Sam Lee 2010
Copyright by Young-Sam Lee 2010 The Dissertation Committee for Young-Sam Lee Certifies that this is the approved version of the following dissertation: Structural and Functional Studies of the Human Mitochondrial DNA Polymerase Committee: Whitney Yin, Supervisor Ian Molineux Kenneth Johnson Tanya Paull Jon Robertus Structural and Functional Studies of the Human Mitochondrial DNA Polymerase by Young-Sam Lee, B.S, M.S. Dissertation Presented to the Faculty of the Graduate School of The University of Texas at Austin in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy The University of Texas at Austin August, 2010 Dedication For my wife, In-Sook Jung. Acknowledgements I would like to appreciate Dr. Whitney Yin for giving me chance to working in her lab and mentoring me through my graduate program. Not only the scientific insights, also the warmness that she gave me and my family encouraged me to pursue my Ph. D. degree in the foreign country. I also would like to thank “a guru of molecular biology” Dr. Ian Molineux and “a guru of enzyme kinetics” Dr. Kenneth Johnson. Without their critical advice, I would not be accomplished my publication. I hope to be a respectable expert in my research field like them. I also should remember friendship and generosity given by many current and former Yin lab members: Hey-Ryung Chang, Qingchao “Eric” Meng, Xu Yang, Jeff Knight, Dr. Michio Matsunaga, Dr. He “River” Quan, Taewung Lee, Xin “Ella” Wang, Jamila Momand, and Max Shay. Most of all, I really appreciate my parents for their endless love and support, and my wife, In-Sook Jung, and my son, Jason Seung-Hyeon Lee who always stand by me with patients during my graduate carrier. -

Introduction of Human Telomerase Reverse Transcriptase to Normal Human Fibroblasts Enhances DNA Repair Capacity
Vol. 10, 2551–2560, April 1, 2004 Clinical Cancer Research 2551 Introduction of Human Telomerase Reverse Transcriptase to Normal Human Fibroblasts Enhances DNA Repair Capacity Ki-Hyuk Shin,1 Mo K. Kang,1 Erica Dicterow,1 INTRODUCTION Ayako Kameta,1 Marcel A. Baluda,1 and Telomerase, which consists of the catalytic protein subunit, No-Hee Park1,2 human telomerase reverse transcriptase (hTERT), the RNA component of telomerase (hTR), and several associated pro- 1School of Dentistry and 2Jonsson Comprehensive Cancer Center, University of California, Los Angeles, California teins, has been primarily associated with maintaining the integ- rity of cellular DNA telomeres in normal cells (1, 2). Telomer- ase activity is correlated with the expression of hTERT, but not ABSTRACT with that of hTR (3, 4). Purpose: From numerous reports on proteins involved The involvement of DNA repair proteins in telomere main- in DNA repair and telomere maintenance that physically tenance has been well documented (5–8). In eukaryotic cells, associate with human telomerase reverse transcriptase nonhomologous end-joining requires a DNA ligase and the (hTERT), we inferred that hTERT/telomerase might play a DNA-activated protein kinase, which is recruited to the DNA role in DNA repair. We investigated this possibility in nor- ends by the DNA-binding protein Ku. Ku binds to hTERT mal human oral fibroblasts (NHOF) with and without ec- without the need for telomeric DNA or hTR (9), binds the topic expression of hTERT/telomerase. telomere repeat-binding proteins TRF1 (10) and TRF2 (11), and Experimental Design: To study the effect of hTERT/ is thought to regulate the access of telomerase to telomere DNA telomerase on DNA repair, we examined the mutation fre- ends (12, 13). -

T-Loops and the Origin of Telomeres E
PERSPECTIVES 66. Steinman, R. M., Mellman, I. S., Muller, W. A. & Cohn, Z. A. Acknowledgments eukaryotes evolved. The presence of t-loops at Endocytosis and the recycling of plasma membrane. H.S. is supported by the Norwegian Cancer Society and the J. Cell Biol. 96, 1–27 (1983). Research Council of Norway, and J.G. by the Swiss National present-day telomeres and their association 67. Lafont, F., Lecat, S., Verkade, P. & Simons, K. Annexin Science Foundation and the Human Frontier Science Programme with proteins that have evolved from RDR XIIIb associates with lipid microdomains to function in Organization. apical delivery. J. Cell Biol. 142, 1413–1427 (1998). factors might be remnants of the original 68. Aniento, F., Gu, F., Parton, R. & Gruenberg, J. Competing interests statement telomere system. Furthermore, the relative An endosomal βcop is involved in the pH-dependent The authors declare that they have no competing financial interests. formation of transport vesicles destined for late ease with which many eukaryotes can main- endosomes. J. Cell Biol. 133, 29–41 (1996). tain telomeres without telomerase might 69. Gu, F., Aniento, F., Parton, R. & Gruenberg, J. Functional Online links dissection of COP-I subunits in the biogenesis of reflect this ancient system of chromosome-end multivesicular endosomes. J. Cell Biol. 139, 1183–1195 DATABASES replication. This proposal ends with a dis- (1997). The following terms in this article are linked online to: 70. Gu, F. & Gruenberg, J. ARF1 regulates pH-dependent Interpro: http://www.ebi.ac.uk/interpro/ cussion of the advantages of the telom- COP functions in the early endocytic pathway. -

PURIFIED THERMOSTABLE NUCLEIC ACID POLYMERASE ENZYME from $I(TERMOTOGA MARITIMA)
Europäisches Patentamt *EP000544789B1* (19) European Patent Office Office européen des brevets (11) EP 0 544 789 B1 (12) EUROPEAN PATENT SPECIFICATION (45) Date of publication and mention (51) Int Cl.7: C12N 15/54, C12N 9/12 of the grant of the patent: 05.03.2003 Bulletin 2003/10 (86) International application number: PCT/US91/05753 (21) Application number: 91915802.2 (87) International publication number: (22) Date of filing: 13.08.1991 WO 92/003556 (05.03.1992 Gazette 1992/06) (54) PURIFIED THERMOSTABLE NUCLEIC ACID POLYMERASE ENZYME FROM $i(TERMOTOGA MARITIMA) GEREINIGTES THERMOSTABILES NUKLEINSÄURE-POLYMERASEENZYM AUS THERMOTOGA MARITIMA ENZYME D’ACIDE NUCLEIQUE THERMOSTABLE PURIFIEE PROVENANT DE L’EUBACTERIE $i(THERMOTOGA MARITIMA) (84) Designated Contracting States: (74) Representative: Poredda, Andreas et al AT BE CH DE DK ES FR GB GR IT LI LU NL SE Roche Diagnostics GmbH, Patentabteilung, (30) Priority: 13.08.1990 US 567244 Sandhofer Strasse 116 68305 Mannheim (DE) (43) Date of publication of application: 09.06.1993 Bulletin 1993/23 (56) References cited: • CHEMICAL ABSTRACTS, vol. 105, no. 5, 04 (73) Proprietor: F. HOFFMANN-LA ROCHE AG August 1986, Columbus, OH (US); R. HUBER et 4002 Basel (CH) al., p. 386, AN 38901u • JOURNAL OF BIOLOGICAL CHEMISTRY, vol. (72) Inventors: 264, no. 11, 15 April 1989, American Society for • GELFAND, David, H. Biochemistry & Molecular Biology Inc., Oakland, CA 94611 (US) Baltimore, MD (US); F.C. LAWYER et al., pp. • LAWYER, Frances, C. 6427-6437 Oakland, CA 94611 (US) • STOFFEL, Susanne El Cerrito, CA 94530 (US) Note: Within nine months from the publication of the mention of the grant of the European patent, any person may give notice to the European Patent Office of opposition to the European patent granted. -

Glucokinase Regulatory Protein Is Essential for the Proper Subcellular Localisation of Liver Glucokinase
FEBS Letters 456 (1999) 332^338 FEBS 22420 Glucokinase regulatory protein is essential for the proper subcellular localisation of liver glucokinase Nu¨ria de la Iglesiaa, Maria Veiga-da-Cunhab, Emile Van Schaftingenb, Joan J. Guinovarta, Juan C. Ferrera;* aDepartament de Bioqu|¨mica i Biologia Molecular, Universitat de Barcelona, Mart|¨ i Franque©s, 1, E-08028 Barcelona, Spain bLaboratory of Physiological Chemistry, Christian de Duve Institute of Cellular Pathology and Universite¨ Catholique de Louvain, B-1200 Brussels, Belgium Received in revised form 24 June 1999 pressed by fructose 1-phosphate, both of which bind to Abstract Glucokinase (GK), a key enzyme in the glucose homeostatic responses of the liver, changes its intracellular GKRP and modify its a¤nity for GK [4]. This 68 kDa protein localisation depending on the metabolic status of the cell. Rat is only found in the livers of species that express GK and liver GK and Xenopus laevis GK, fused to the green fluorescent although there is some evidence for its presence in pancreatic protein (GFP), concentrated in the nucleus of cultured rat tissue [5,6], a direct demonstration is still not available. hepatocytes at low glucose and translocated to the cytoplasm at In in vitro assays, rat GKRP can inhibit human pancreatic high glucose. Three mutant forms of Xenopus GK with reduced GK, which shows a high degree of identity to the rat liver affinity for GK regulatory protein (GKRP) did not concentrate isoform [7], as well as Xenopus laevis liver GK [8], a more in the hepatocyte nuclei, even at low glucose. In COS-1 and distantly related protein. -

Telomeres.Pdf
Telomeres Secondary article Elizabeth H Blackburn, University of California, San Francisco, California, USA Article Contents . Introduction Telomeres are specialized DNA–protein structures that occur at the ends of eukaryotic . The Replication Paradox chromosomes. A special ribonucleoprotein enzyme called telomerase is required for the . Structure of Telomeres synthesis and maintenance of telomeric DNA. Synthesis of Telomeric DNA by Telomerase . Functions of Telomeres Introduction . Telomere Homeostasis . Alternatives to Telomerase-generated Telomeric DNA Telomeres are the specialized chromosomal DNA–protein . Evolution of Telomeres and Telomerase structures that comprise the terminal regions of eukaryotic chromosomes. As discovered through studies of maize and somes. One critical part of this protective function is to fruitfly chromosomes in the 1930s, they are required to provide a means by which the linear chromosomal DNA protect and stabilize the genetic material carried by can be replicated completely, without the loss of terminal eukaryotic chromosomes. Telomeres are dynamic struc- DNA nucleotides from the 5’ end of each strand of this tures, with their terminal DNA being constantly built up DNA. This is necessary to prevent progressive loss of and degraded as dividing cells replicate their chromo- terminal DNA sequences in successive cycles of chromo- somes. One strand of the telomeric DNA is synthesized by somal replication. a specialized ribonucleoprotein reverse transcriptase called telomerase. Telomerase is required for both -

DNA Replication
9 DNA Replication To understand the chemistry Living cells must be able to duplicate their entire set of genetic instructions Goal of DNA synthesis and how every time they divide. Likewise, multicellular organisms must be able to DNA is replicated with high pass on complete copies of their genetic information to future generations. accuracy. This requires that the instructions are stored in a form that is capable of Objectives being duplicated. As we have seen (and will return to in Chapter 11), genetic instructions are embedded in the order of nucleobases in the DNA. As we After this chapter, you should be able to have also seen, the self-complementary nature of the double helix provides • describe the experiment that proved a simple (in principle) templating mechanism for replicating DNA into two that DNA replication is semi- identical copies. Only DNA (and in some instances RNA when, as in the conservative. case of the genomes of RNA viruses, it is used as a repository of genetic • diagram the reaction for information) are self-complementary and hence capable of being replicated. phosphodiester bond formation. The other three categories of macromolecules—carbohydrates, lipids, and • explain the energetics of DNA proteins—are not self-complementary and do not serve as templates for synthesis. their own production. Instead, the synthesis of these macromolecules is • explain why the 5’-to-3’ rule creates a ultimately directed by information stored in DNA through the action of conundrum during replication. enzymes and other proteins. Here we focus on the chemical and enzymatic • explain how DNA is replicated mechanisms by which DNA acts as a template for its own duplication and accurately. -

Getting Ready for DNA Duplication
INSIGHT INTRACELLULAR ORGANIZATION Getting ready for DNA duplication The discovery of a biomolecular condensate involved in DNA replication has wide-ranging implications. NINA Y YAO AND MICHAEL E O’DONNELL involves regions with different physical proper- Related research article Parker MW, Bell ties separating from each other). The molecules M, Mir M, Kao JA, Darzacq X, Botchan MR, required for the formation of condensates are Berger JM. 2019. A new class of disordered called scaffolding factors, while the molecules elements controls DNA replication through encapsulated inside are known as clients. A initiator self-assembly. eLife 8:e48562. DOI: given condensate typically contains only those 10.7554/eLife.48562 clients involved in the relevant reaction. Now, in eLife, Michael Botchan (UC Berkeley), James Berger (Johns Hopkins) and colleagues – including Matthew Parker as first author – report on the discovery of a biomolecular condensate esearch into biomolecular condensates – required for the initiation of DNA replication in self-contained regions within cells where the fruit fly Drosophila melanogaster R specific reactions take place – is taking (Parker et al., 2019). Three proteins – ORC, cell biology by storm. Biomolecular condensates Cdc6 and Cdt1 – form the scaffold along with do not have membranes, but they are able to DNA, while a hexamer ring protein called keep the proteins and/or nucleic acids involved Mcm2-7 is the client. During the G1 phase of the in a particular reaction separate from the rest of cell cycle, the scaffold proteins load two Mcm2- the cell. Biomolecular condensates do not have 7 hexamers onto the DNA to form the pre-repli- membranes, but they are able to keep the mole- cative complex, which marks the spot where cules involved in a particular reaction (usually DNA replication will begin (Figure 1A–C; proteins, but sometimes also nucleic acids) sepa- Bell and Labib, 2016; Bleichert et al., 2017). -

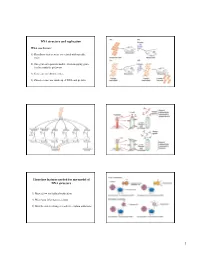
Arthur Kornberg Discovered (The First) DNA Polymerase Four
Arthur Kornberg discovered (the first) DNA polymerase Using an “in vitro” system for DNA polymerase activity: 1. Grow E. coli 2. Break open cells 3. Prepare soluble extract 4. Fractionate extract to resolve different proteins from each other; repeat; repeat 5. Search for DNA polymerase activity using an biochemical assay: incorporate radioactive building blocks into DNA chains Four requirements of DNA-templated (DNA-dependent) DNA polymerases • single-stranded template • deoxyribonucleotides with 5’ triphosphate (dNTPs) • magnesium ions • annealed primer with 3’ OH Synthesis ONLY occurs in the 5’-3’ direction Fig 4-1 E. coli DNA polymerase I 5’-3’ polymerase activity Primer has a 3’-OH Incoming dNTP has a 5’ triphosphate Pyrophosphate (PP) is lost when dNMP adds to the chain E. coli DNA polymerase I: 3 separable enzyme activities in 3 protein domains 5’-3’ polymerase + 3’-5’ exonuclease = Klenow fragment N C 5’-3’ exonuclease Fig 4-3 E. coli DNA polymerase I 3’-5’ exonuclease Opposite polarity compared to polymerase: polymerase activity must stop to allow 3’-5’ exonuclease activity No dNTP can be re-made in reversed 3’-5’ direction: dNMP released by hydrolysis of phosphodiester backboneFig 4-4 Proof-reading (editing) of misincorporated 3’ dNMP by the 3’-5’ exonuclease Fidelity is accuracy of template-cognate dNTP selection. It depends on the polymerase active site structure and the balance of competing polymerase and exonuclease activities. A mismatch disfavors extension and favors the exonuclease.Fig 4-5 Superimposed structure of the Klenow fragment of DNA pol I with two different DNAs “Fingers” “Thumb” “Palm” red/orange helix: 3’ in red is elongating blue/cyan helix: 3’ in blue is getting edited Fig 4-6 E. -

DNA POLYMERASE III HOLOENZYME: Structure and Function of a Chromosomal Replicating Machine
Annu. Rev. Biochem. 1995.64:171-200 Copyright Ii) 1995 byAnnual Reviews Inc. All rights reserved DNA POLYMERASE III HOLOENZYME: Structure and Function of a Chromosomal Replicating Machine Zvi Kelman and Mike O'Donnell} Microbiology Department and Hearst Research Foundation. Cornell University Medical College. 1300York Avenue. New York. NY }0021 KEY WORDS: DNA replication. multis ubuni t complexes. protein-DNA interaction. DNA-de penden t ATPase . DNA sliding clamps CONTENTS INTRODUCTION........................................................ 172 THE HOLO EN ZYM E PARTICL E. .......................................... 173 THE CORE POLYMERASE ............................................... 175 THE � DNA SLIDING CLAM P............... ... ......... .................. 176 THE yC OMPLEX MATCHMAKER......................................... 179 Role of ATP . .... .............. ...... ......... ..... ............ ... 179 Interaction of y Complex with SSB Protein .................. ............... 181 Meclwnism of the yComplex Clamp Loader ................................ 181 Access provided by Rockefeller University on 08/07/15. For personal use only. THE 't SUBUNIT . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. 182 Annu. Rev. Biochem. 1995.64:171-200. Downloaded from www.annualreviews.org AS YMMETRIC STRUC TURE OF HOLO EN ZYM E . 182 DNA PO LYM ER AS E III HOLO ENZ YME AS A REPLIC ATING MACHINE ....... 186 Exclwnge of � from yComplex to Core .................................... 186 Cycling of Holoenzyme on the LaggingStrand -

Teacher Notes Science Nspired
DNA Replication TEACHER NOTES SCIENCE NSPIRED Science Objectives In this lesson, students will • Explore how the structure of DNA supports its semi-conservative replication • Identify the name and function of several key enzymes in DNA replication • Recognize the function of Okazaki fragments in DNA replication Vocabulary TI-Nspire™ Technology Skills: • Double helix • Leading strand • Download a TI-Nspire • Lagging strand • Polymerase I • Polymerase III • Helicase document • Base pairs • Okazaki Fragments • Open a document • Primase • Move between pages • Open Directions Box About the Lesson • Explore ‘Hot Spots’ • Using three simulations, students will interact with DNA replication to explore semi-conservative replication and identify Tech Tips: specific enzymes and their roles in replication. Assessments are Make sure that students know embedded in the activity to engage discussion and gauge how to move between pages by learning. pressing /¡ (left arrow) and • As a result, students will: /¢ (right arrow). • Learn the basic functions of the following DNA replication enzymes: helicase, primase, ligase, polymerase I and III. Lesson Materials: • Learn how the double helix reproduces in a semi- Student Activity • DNA_Replication_Student.do conservative fashion c • Learn the role and function of Okazaki fragments in • DNA_Replication_Student.pd replication of the lagging strand. f TI-Nspire document TI-Nspire™ Navigator™ • DNA_Replication.tns Not required. If using Navigator: • Send out the DNA_Replication.tns file. • Monitor student progress -
DNA Structure and Replication Three Key Features Needed for Any Model
DNA structure and replication What was known? 1) Hereditary factors were associated with specific traits 2) One-gene-one-protein model - from mapping genes for biosynthetic pathways 3) Genes are on chromosomes 4) Chromosomes are made up of DNA and protein Three key features needed for any model of DNA structure 1) Must allow for faithful replication 2) Must have information content 3) Must be able to change in order to explain mutations 1 Rosalind Franklin James Watson Francis Crick X-ray diffraction Maurice Wilkins Photograph of B-DNA Linus Pauling Nature 171, 737-738 (1953) © Macmillan Publishers Ltd. Molecular structure of Nucleic Acids WATSON, J. D. & CRICK, F. H. C. Features of the double helix A Structure for Deoxyribose Nucleic Acid 1) Two parallel strands We wish to suggest a structure for the salt of deoxyribose nucleic acid (D.N.A.). This structure has novel features which are of considerable biological interest………… 2) Bases held together by H-bonds 3) Phosphodiester backbone 4) The base is attached to the position 1 on the sugar 5) Base pair stack - provides Figure 1. This figure is purely diagrammatic. The two ribbons symbolize the two phophate-sugar chains, and the horizonal rods stability the pairs of bases holding the chains together. The vertical line marks the fibre axis. 6) Contains a major and …………….It has not escaped our notice that the specific pairing we minor groove have postulated immediately suggests a possible copying mechanism for the genetic material. Three key features needed for any model of DNA structure