Federally-Listed Plants
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Butte Co. Meadowfoam
U.S. Fish and Wildlife Service, and the U.S. Air Force, respectively. All of the Fort Ord occurrences are on land within the Habitat Management Plan Habitat Reserve lands and will be conserved and managed in perpetuity (W. Collins in litt. 2005; U.S. Army Corps of Engineers 1997). The population at Travis Air Force Base, including over 20 acres of adjacent restored vernal pools, is protected as a special ecological preserve, with protective measures and appropriate management for the species provided in the Travis Air Force Base Land Management Plan. Seasonal managed cattle grazing has been returned to two conservation sites supporting Lasthenia conjugens: 1) the Warm Springs Seasonal Wetland Unit of the Don Edwards San Francisco Bay National Wildlife Refuge in Alameda County, and 2) the State Route 4 Preserve managed by the Muir Heritage Land Trust in Contra Costa County. The L. conjugens population at the Warm Springs Unit has declined during the last 10 years due to many factors including competition by nonnative plant species. During this time period, grazing, which occurred intermittently at the Warm Springs Unit since the 1800s, has been excluded by the Refuge until a management plan could be developed. The decline in the L. conjugens population at the Warm Springs Unit cannot be attributed to a single factor, but most likely results from the complex interaction of several variables including current and historical land uses, the abiotic environment, and annual climatic variation. The increasing dominance of nonnative grasses, however, coincides with the suspension of livestock grazing, suggesting that the lack of a disturbance regime may be a primary factor in the degradation of habitat for L. -

Loch Lomond Button-Celery)
Natural Diversity Data Base 2003). The Sacramento National Wildlife Refuge populations have been monitored annually since 1992 (J. Silveira in litt. 2000). One additional occurrence of C. hooveri in Merced County is on private land (the Bert Crane Ranch) that is protected from development by a conservation easement (J. Silveira in litt. 2000). We funded a status survey for Chamaesyce hooveri and other vernal pool plants in 1986 and 1987 (Stone et al. 1988), resulting in 10 new occurrences. We and the California Department of Fish and Game jointly funded an ecological study of the Vina Plains Preserve pools, which was conducted by faculty from California State University, Chico (Alexander and Schlising 1997). Independent surveys conducted by Joseph Silveira led to discovery of the Merced and Glenn county occurrences (J. Silveira in litt. 2000). Private landowners also have contributed to conservation of this species. One pool in Tehama County was fenced by the property owner in the late 1980s, to exclude livestock (Stone et al. 1988). 3. ERYNGIUM CONSTANCEI (LOCH LOMOND BUTTON-CELERY) a. Description and Taxonomy Taxonomy.—Loch Lomond button-celery, specifically known as Eryngium constancei (Sheikh 1983), is a member of the carrot family (Apiaceae). This species was only recently described and therefore has no history of name changes. The common name was derived from the type locality, Loch Lomond, which is in Lake County (Sheikh 1983). Other common names for this species are Loch Lomond coyote-thistle (Skinner and Pavlik 1994) and Constance’s coyote-thistle (Smith et al. 1980). Description and Identification.—Certain features are common to species of the genus Eryngium. -

Recovery Plan for the Santa Rosa Plain
U.S. Fish & Wildlife Service Recovery Plan for the Santa Rosa Plain Blennosperma bakeri (Sonoma sunshine) Lasthenia burkei (Burke’s goldfields) Limnanthes vinculans (Sebastopol meadowfoam) California tiger salamander Sonoma County Distinct Population Segment (Ambystoma californiense) Lasthenia burkei Blennosperma bakeri Limnanthes vinculans Jo-Ann Ordano J. E. (Jed) and Bonnie McClellan Jo-Ann Ordano © 2004 California Academy of Sciences © 1999 California Academy of Sciences © 2005 California Academy of Sciences Sonoma County California Tiger Salamander Gerald Corsi and Buff Corsi © 1999 California Academy of Sciences Disclaimer Recovery plans delineate reasonable actions that are believed to be required to recover and/or protect listed species. We, the U.S. Fish and Wildlife Service, publish recovery plans, sometimes preparing them with the assistance of recovery teams, contractors, state agencies, Tribal agencies, and other affected and interested parties. Objectives will be attained and any necessary funds made available subject to budgetary and other constraints affecting the parties involved, as well as the need to address other priorities. Costs indicated for action implementation and time of recovery are estimates and subject to change. Recovery plans do not obligate other parties to undertake specific actions, and may not represent the views or the official positions of any individuals or agencies involved in recovery plan formulation, other than the Service. Recovery plans represent our official position only after they have been signed by the Director or Regional Director as approved. Approved recovery plans are subject to modification as dictated by new findings, changes in species status, and the completion of recovery actions. LITERATURE CITATION SHOULD READ AS FOLLOWS: U.S. -

4.4 BIOLOGICAL RESOURCES 4.4.1 Regulatory Setting
Ascent Environmental Administrative Draft – For Internal Review and Deliberation Biological Resources 4.4 BIOLOGICAL RESOURCES This section describes the potential effects of the project on biological resources. This section also addresses biological resources known or with potential to occur in the project vicinity, including common vegetation and habitat types, sensitive plant communities, and special-status plant and animal species. The analysis includes a description of the existing environmental conditions, the methods used for assessment, the potential direct and indirect impacts of project implementation not included in the 2005 Subsequent Environmental Impact Report (SEIR) for the Hay Road Landfill Project (Solano County 2005), and mitigation measures recommended to address impacts determined to be significant or potentially significant. The data and documents reviewed in preparation of this analysis included: 2005 SEIR for the Hay Road Landfill Project (Solano County 2005); Burrowing Owl Habitat Assessment (ESA 2016a); California Tiger Salamander Habitat Assessment (ESA 2016b); Branchiopod Survey Report (ESA 2016c); Special-status Plant Survey Report (ESA 2016d); Organics Transload Facility Habitat Assessment (ESA 2017a); Hydro Flow Analysis (ESA 2017b); Contra Costa Goldfields Survey Report (ESA 2017c); Delta Green Ground Beetle Survey Report and Supplemental Habitat Assessment Report (Entomological Consulting Services, Ltd. 2016, 2018); reconnaissance-level survey of the project site conducted on August 7, 2017; records search and GIS query of the California Natural Diversity Database (CNDDB) within 5 miles of the project site (2018); California Native Plant Society (CNPS), Rare Plant Program database search of the Allendale, Dixon, Saxon, Elmira, Dozier, Liberty Island, Denverton, Birds Landing, and Rio Vista U.S. Geological Service 7.5-minute quadrangles (CNPS 2018); eBird online database of bird observations (eBird 2018); and aerial photographs of the project site and surrounding area. -

The Biological Resources Section Provides Background Information
4.5 BIOLOGICAL RESOURCES The Biological resources section provides background information on sensitive biological resources within Napa County, the regulations and programs that provide for their protection, and an assessment of the potential impacts to biological resources of implementing the Napa County General Plan Update. This section is based upon information presented in the Biological Resources Chapter of the Napa County Baseline Data Report (Napa County, BDR 2005). Additional information on the topics presented herein can be found in these documents. Both documents are incorporated into this section by reference. This section addresses biological resources other than fisheries which are separately addressed in Section 4.6. 4.5.1 SETTING REGIONAL SETTING The Napa County is located in the Coast Ranges Geomorphic Province. This province is bounded on the west by the Pacific Ocean and on the east by the Great Valley geomorphic province. A dominant characteristic of the Coast Ranges Province is the general northwest- southeast orientation of its valleys and ridgelines. In Napa County, located in the eastern, central section of the province, this trend consists of a series of long, linear, major and lesser valleys, separated by steep, rugged ridge and hill systems of moderate relief that have been deeply incised by their drainage systems. The County is located within the California Floristic Province, the portion of the state west of the Sierra Crest that is known to be particularly rich in endemic plant species (Hickman 1993, Stein et al. 2000). LOCAL SETTING The County’s highest topographic feature is Mount St. Helena, which is located in the northwest corner of the County and whose peak elevation is 4,343 feet. -

MITIGATED NEGATIVE DECLARATION for the Tesla Road
Draft INITIAL STUDY/ MITIGATED NEGATIVE DECLARATION for the Tesla Road Winery ALAMEDA COUNTY CALIFORNIA Prepared for: Alameda County CDA Planning Division Prepared by: Denise Duffy & Associates Contact: Denise Duffy 947 Cass St. Suite 5 Monterey, California 93940 July 31, 2015 This page left intentionally blank. MITIGATED NEGATIVE DECLARATION Project: Tesla Road Winery Lead Agency: Alameda County PROJECT DESCRIPTION This Mitigated Negative Declaration (MND), supported by the attached Initial Study (IS), evaluates the environmental effects of a proposed multi‐use wine facility at the northeast corner of the Greenville Road and Tesla Road intersection outside of Livermore, within unincorporated Alameda County, California. The applicant, RAO Company, is proposing the construction of a new 19,944 square foot (sq. ft.) building on the the property. The building’s primary function would be to provide space for wine tasting, tours, and special events, and administrative offices for employees. The building would provide a dedicated space to process wine, serve customers, and hold events. Alameda County is the lead agency for this project and has prepared this MND. FINDINGS An IS has been prepared to assess the projects potential effects on the environment and the significance of those effects. Based on the Initial Study, it has been determined that the proposed project would not have any significant effects on the environment once mitigation measures are implemented. This conclusion is supported by the following findings: 1. The proposed project would have no impact related to greenhouse gas emissions, aesthetics, agricultural resources, hazards/hazardous materials, land use/planning mineral resources, population/housing, public services, and recreation. -

California Wetlands
VOL. 46, NO.2 FREMONTIA JOURNAL OF THE CALIFORNIA NATIVE PLANT SOCIETY California Wetlands 1 California Native Plant Society CNPS, 2707 K Street, Suite 1; Sacramento, CA 95816-5130 Phone: (916) 447-2677 • Fax: (916) 447-2727 FREMONTIA www.cnps.org • [email protected] VOL. 46, NO. 2, November 2018 Memberships Copyright © 2018 Members receive many benefits, including a subscription toFremontia California Native Plant Society and the CNPS Bulletin. Look for more on inside back cover. ISSN 0092-1793 (print) Mariposa Lily.............................$1,500 Family..............................................$75 ISSN 2572-6870 (online) Benefactor....................................$600 International or library...................$75 Patron............................................$300 Individual................................$45 Gordon Leppig, Editor Plant lover.....................................$100 Student/retired..........................$25 Michael Kauffmann, Editor & Designer Corporate/Organizational 10+ Employees.........................$2,500 4-6 Employees..............................$500 7-10 Employees.........................$1,000 1-3 Employees............................$150 Staff & Contractors Dan Gluesenkamp: Executive Director Elizabeth Kubey: Outreach Coordinator Our mission is to conserve California’s Alfredo Arredondo: Legislative Analyst Sydney Magner: Asst. Vegetation Ecologist native plants and their natural habitats, Christopher Brown: Membership & Sales David Magney: Rare Plant Program Manager and increase understanding, -
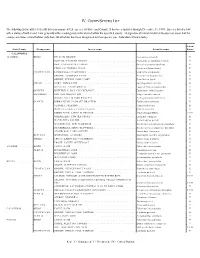
Iv. County/Species List
IV. COUNTY/SPECIES LIST The following list identifies federally listed or proposed U.S. species by State and County. It has been updated through December 31, 1999. Species listed below with a status of both E and T are generally either endangered or threatened within the specified county. Designation of critical habitat (CH) does not mean that the county constitutes critical habitat, only that critical habitat has been designated for that species (see Addendum A Instructions). Action/ State/County Group name Inverse name Scientific name Status CALIFORNIA ALAMEDA ......... BIRDS ....... PELICAN, BROWN ........................................ Pelicanus occidentalis E PLOVER, WESTERN SNOWY ................................ Charadrius alexandrinus nivosus T RAIL, CALIFORNIA CLAPPER ............................... Rallus longirostris obsoletus E TERN, CALIFORNIA LEAST ................................. Sterna antillarum browni E CRUSTACEAN . LINDERIELLA, CALIFORNIA ................................ Linderiella occidentalis E SHRIMP, LONGHORN FAIRY ................................ Branchinecta longiantenna E SHRIMP, VERNAL POOL FAIRY .............................. Branchinecta lynchi T FISHES ...... GOBY, TIDEWATER ...................................... Eucyclogobius newberryi E SPLITTAIL, SACRAMENTO ................................. Pogonichthys macrolepidotus T INSECTS ..... BUTTERFLY, BAY CHECKERSPOT ........................... Euphydryas editha bayensis T MAMMALS ... FOX, SAN JOAQUIN KIT .................................... Vulpes macrotis -

A Checklist of Vascular Plants Endemic to California
Humboldt State University Digital Commons @ Humboldt State University Botanical Studies Open Educational Resources and Data 3-2020 A Checklist of Vascular Plants Endemic to California James P. Smith Jr Humboldt State University, [email protected] Follow this and additional works at: https://digitalcommons.humboldt.edu/botany_jps Part of the Botany Commons Recommended Citation Smith, James P. Jr, "A Checklist of Vascular Plants Endemic to California" (2020). Botanical Studies. 42. https://digitalcommons.humboldt.edu/botany_jps/42 This Flora of California is brought to you for free and open access by the Open Educational Resources and Data at Digital Commons @ Humboldt State University. It has been accepted for inclusion in Botanical Studies by an authorized administrator of Digital Commons @ Humboldt State University. For more information, please contact [email protected]. A LIST OF THE VASCULAR PLANTS ENDEMIC TO CALIFORNIA Compiled By James P. Smith, Jr. Professor Emeritus of Botany Department of Biological Sciences Humboldt State University Arcata, California 13 February 2020 CONTENTS Willis Jepson (1923-1925) recognized that the assemblage of plants that characterized our flora excludes the desert province of southwest California Introduction. 1 and extends beyond its political boundaries to include An Overview. 2 southwestern Oregon, a small portion of western Endemic Genera . 2 Nevada, and the northern portion of Baja California, Almost Endemic Genera . 3 Mexico. This expanded region became known as the California Floristic Province (CFP). Keep in mind that List of Endemic Plants . 4 not all plants endemic to California lie within the CFP Plants Endemic to a Single County or Island 24 and others that are endemic to the CFP are not County and Channel Island Abbreviations . -

A Classification of the Chloridoideae (Poaceae)
Molecular Phylogenetics and Evolution 55 (2010) 580–598 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev A classification of the Chloridoideae (Poaceae) based on multi-gene phylogenetic trees Paul M. Peterson a,*, Konstantin Romaschenko a,b, Gabriel Johnson c a Department of Botany, National Museum of Natural History, Smithsonian Institution, Washington, DC 20013, USA b Botanic Institute of Barcelona (CSICÀICUB), Pg. del Migdia, s.n., 08038 Barcelona, Spain c Department of Botany and Laboratories of Analytical Biology, Smithsonian Institution, Suitland, MD 20746, USA article info abstract Article history: We conducted a molecular phylogenetic study of the subfamily Chloridoideae using six plastid DNA Received 29 July 2009 sequences (ndhA intron, ndhF, rps16-trnK, rps16 intron, rps3, and rpl32-trnL) and a single nuclear ITS Revised 31 December 2009 DNA sequence. Our large original data set includes 246 species (17.3%) representing 95 genera (66%) Accepted 19 January 2010 of the grasses currently placed in the Chloridoideae. The maximum likelihood and Bayesian analysis of Available online 22 January 2010 DNA sequences provides strong support for the monophyly of the Chloridoideae; followed by, in order of divergence: a Triraphideae clade with Neyraudia sister to Triraphis; an Eragrostideae clade with the Keywords: Cotteinae (includes Cottea and Enneapogon) sister to the Uniolinae (includes Entoplocamia, Tetrachne, Biogeography and Uniola), and a terminal Eragrostidinae clade of Ectrosia, Harpachne, and Psammagrostis embedded Classification Chloridoideae in a polyphyletic Eragrostis; a Zoysieae clade with Urochondra sister to a Zoysiinae (Zoysia) clade, and a Grasses terminal Sporobolinae clade that includes Spartina, Calamovilfa, Pogoneura, and Crypsis embedded in a Molecular systematics polyphyletic Sporobolus; and a very large terminal Cynodonteae clade that includes 13 monophyletic sub- Phylogenetic trees tribes. -

4.4 Biological Resources
4.4 BIOLOGICAL RESOURCES This section discusses biological resources in Vacaville and evaluates the potential biological re- source impacts resulting from the spatial location of development that would be allowed by the proposed General Plan and Energy and Conservation Action Strategy (ECAS). The following evaluation assesses special-status species, sensitive biological communities, wetlands, migratory species, and policies and plans intended to protect biological resources. As noted in Chapter 3, Project Description, impacts are determined by comparing the proposed General Plan and ECAS to existing conditions, rather than to the existing General Plan. The following evaluation is based on a spatial analysis and examines the effects of the location of potential development on biological resources and identifies conflicts with plans, policies, regulations, and ordinances relating to biological resources. A. Regulatory Framework 1. Federal Laws and Regulations This section summarizes federal laws and regulations that apply to biological resources within the EIR Study Area. a. Section 404 of the Clean Water Act The US Army Corps Engineers (Corps), the federal agency charged with investigating, develop- ing, and maintaining the country’s water and related resources, is responsible under Section 404 of the Clean Water Act for regulating the discharge of fill material into waters of the United States. Waters of the United States and their lateral limits are defined in Part 328.3(a) of Title 33 of the Code of Federal Regulations (CFR) and include streams that are tributaries to navigable waters and adjacent wetlands. The lateral limits of jurisdiction for a non-tidal stream are meas- ured at the line of the Ordinary High Water Mark1 or the limit of adjacent wetlands.2 Any per- manent extension of the limits of an existing water of the United States, whether natural or hu- man-made, results in a similar extension of Corps jurisdiction.3 Waters of the United States fall into two broad categories: wetlands and other waters. -

Status and Distribution of Contra Costa Goldfields in Solano County, California
THE STATUS AND DISTRIBUTION OF CONTRA COSTA GOLDFIELDS IN SOLANO COUNTY, CALIFORNIA RESULTS OF CONTRA COSTA GOLDFIELD POPULATION MONITORING FOR 2006, 2007, 2008, AND 2009 SOLANO COUNTY, CALIFORNIA June 30, 2010 THE STATUS AND DISTRIBUTION OF CONTRA COSTA GOLDFIELDS IN SOLANO COUNTY, CALIFORNIA RESULTS OF CONTRA COSTA GOLDFIELD MONITORING, 2006-2009 SOLANO COUNTY, CALIFORNIA Submitted to: Solano County Water Agency Prepared by: LSA Associates, Inc. 157 Park Place Point Richmond, California 94801 (510) 236-6810 LSA Project No. SCD430, SCD0601, SWG0701, SWG0801, and SWG0901 June 30, 2010 LSA ASSOCIATES, INC. CONTRA COSTA GOLDFIELD POPULATION ASSESSMENT JUNE 2010 SOLANO COUNTY WATER AGENCY TABLE OF CONTENTS INTRODUCTION ........................................................................................................................................4 CONTRA COSTA GOLDFIELDS....................................................................................................5 GENERAL SITE CHARACTERISTICS...........................................................................................6 Barnfield...................................................................................................................................6 Director’s Guild .......................................................................................................................7 Goldfield Conservation Bank...................................................................................................7 Jehovah’s Witness Complex ....................................................................................................8