Fish Community Ecology in an Altered River Delta: Spatial Patterns in Species Composition, Life History Strategies, and Biomass
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
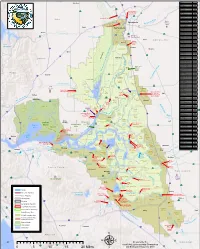
0 5 10 15 20 Miles Μ and Statewide Resources Office
Woodland RD Name RD Number Atlas Tract 2126 5 !"#$ Bacon Island 2028 !"#$80 Bethel Island BIMID Bishop Tract 2042 16 ·|}þ Bixler Tract 2121 Lovdal Boggs Tract 0404 ·|}þ113 District Sacramento River at I Street Bridge Bouldin Island 0756 80 Gaging Station )*+,- Brack Tract 2033 Bradford Island 2059 ·|}þ160 Brannan-Andrus BALMD Lovdal 50 Byron Tract 0800 Sacramento Weir District ¤£ r Cache Haas Area 2098 Y o l o ive Canal Ranch 2086 R Mather Can-Can/Greenhead 2139 Sacramento ican mer Air Force Chadbourne 2034 A Base Coney Island 2117 Port of Dead Horse Island 2111 Sacramento ¤£50 Davis !"#$80 Denverton Slough 2134 West Sacramento Drexler Tract Drexler Dutch Slough 2137 West Egbert Tract 0536 Winters Sacramento Ehrheardt Club 0813 Putah Creek ·|}þ160 ·|}þ16 Empire Tract 2029 ·|}þ84 Fabian Tract 0773 Sacramento Fay Island 2113 ·|}þ128 South Fork Putah Creek Executive Airport Frost Lake 2129 haven s Lake Green d n Glanville 1002 a l r Florin e h Glide District 0765 t S a c r a m e n t o e N Glide EBMUD Grand Island 0003 District Pocket Freeport Grizzly West 2136 Lake Intake Hastings Tract 2060 l Holland Tract 2025 Berryessa e n Holt Station 2116 n Freeport 505 h Honker Bay 2130 %&'( a g strict Elk Grove u Lisbon Di Hotchkiss Tract 0799 h lo S C Jersey Island 0830 Babe l Dixon p s i Kasson District 2085 s h a King Island 2044 S p Libby Mcneil 0369 y r !"#$5 ·|}þ99 B e !"#$80 t Liberty Island 2093 o l a Lisbon District 0307 o Clarksburg Y W l a Little Egbert Tract 2084 S o l a n o n p a r C Little Holland Tract 2120 e in e a e M Little Mandeville -

Transitions for the Delta Economy
Transitions for the Delta Economy January 2012 Josué Medellín-Azuara, Ellen Hanak, Richard Howitt, and Jay Lund with research support from Molly Ferrell, Katherine Kramer, Michelle Lent, Davin Reed, and Elizabeth Stryjewski Supported with funding from the Watershed Sciences Center, University of California, Davis Summary The Sacramento-San Joaquin Delta consists of some 737,000 acres of low-lying lands and channels at the confluence of the Sacramento and San Joaquin Rivers (Figure S1). This region lies at the very heart of California’s water policy debates, transporting vast flows of water from northern and eastern California to farming and population centers in the western and southern parts of the state. This critical water supply system is threatened by the likelihood that a large earthquake or other natural disaster could inflict catastrophic damage on its fragile levees, sending salt water toward the pumps at its southern edge. In another area of concern, water exports are currently under restriction while regulators and the courts seek to improve conditions for imperiled native fish. Leading policy proposals to address these issues include improvements in land and water management to benefit native species, and the development of a “dual conveyance” system for water exports, in which a new seismically resistant canal or tunnel would convey a portion of water supplies under or around the Delta instead of through the Delta’s channels. This focus on the Delta has caused considerable concern within the Delta itself, where residents and local governments have worried that changes in water supply and environmental management could harm the region’s economy and residents. -

Programmatic Environmental Impact Report
Water Hyacinth Control Program FINAL Programmatic Environmental Impact Report Volume I – Chapters 1 to 7 November 30, 2009 A program for effective control of Water Hyacinth in the Sacramento-San Joaquin Delta and its tributaries. Copies of this Final Programmatic Environmental Impact Report in hard copy form, or on computer compact disc (CD), can be obtained from the California Department of Boating and Waterways. To request a report copy, please contact: Ms. Terri Ely Aquatic Weed Program California Department of Boating and Waterways 2000 Evergreen Street, Suite 100 Sacramento, California 95815 (916) 263-8138 [email protected] Cover photo: March 14, 2008, by NewPoint Group, Inc., of the Wheeler Island Duck Club, at Honker Bay. [PARTIAL] Water Hyacinth Control Program Water Hyacinth Control Program A program for effective control of Water Hyacinth in the Sacramento-San Joaquin Delta and its tributaries. FINAL Programmatic Environmental Impact Report Volume I – Chapters 1 to 7 November 30, 2009 Prepared by: The California Department of Boating and Waterways With Technical Assistance from: NewPoint Group, Inc. 2555 Third Street, Suite 215 Sacramento, California 95818 (916) 442-0508 www.newpointgroup.com ~----Pei:at f~m.A; _ _,__,..._... AniJru--~- ' --sepat Table of Contents Volume I – Chapters 1 to 7 Page Acronyms and Abbreviations ......................................................... AA-1 Executive Summary.......................................................................... ES-1 1. Introduction ........................................................................................ -
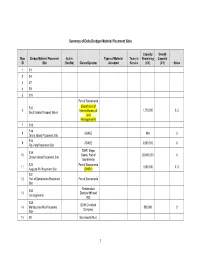
Summary of Delta Dredged Material Placement Sites
Summary of Delta Dredged Material Placement Sites Capacity Overall Map Dredge Material Placement Active Types of Material Years in Remaining Capacity ID Site (Yes/No) Owner/Operator Accepted Service (CY) (CY) Notes 1 S1 2 S4 3 S7 4 S9 5 S11 Port of Sacramento S12 (Department of 6 1,710,000 3, 5 South Island Prospect Island Interior Bureau of Land Management?) 7 S13 S14 8 USACE N/A 3 Grand Island Placement Site S16 9 USACE 3,000,000 3 Rio Vista Placement Site DWR, Mega S19 10 Sands, Port of 20,000,000 3 Decker Island Placement Site Sacramento S20 Port of Sacramento 11 1,000,000 3, 5 Augusta Pit Placement Site (DWR?) S31 12 Port of Sacramento Placement Port of Sacramento Site Reclamation S32 13 Districts 999 and (six segments) 900 S35 DOW Chemical 14 Montezuma Hills Placement 890,000 3 Company Site 15 SX Sacramento Muni 1 Capacity Overall Map Dredge Material Placement Active Types of Material Years in Remaining Capacity ID Site (Yes/No) Owner/Operator Accepted Service (CY) (CY) Notes Utility District Sherman Lake (Sherman 16 USACE 3,000,000 3 Island?) 17 Montezuma Wetlands Project Montezuma LLC Montezuma Wetlands 18 Montezuma LLC Rehandling Site Expanded Scour Pond Dredge material 19 Placement Site (also called Yes DWR according to WDR #R5- 250,000 1, 2, 3,4 Sherman Island?) 2004-0061 Port of Stockton McCormack Pit Placement maintenance material 20 Site (also called Sherman Yes DWR only 250,000 3,4 Island?) WDR R5-2003-0145 Proposed Iron House Levy repair and 21 Jersey Island Placement Site Restoration 3 Sanitation District maintenance -

Pipeline Installation by the Hole Intersect Method to Protect Facilities Crossing Under Navigable Waterways
PipelinePipeline InstallationInstallation byby thethe HoleHole IntersectIntersect MethodMethod toto ProtectProtect FacilitiesFacilities CrossingCrossing UnderUnder NavigableNavigable WaterwaysWaterways • L-57A (18”) installed 1949 by Standard Oil. Floated during flooding of Mildred Island, and is partially decommissioned. • L-57B (22”) installed 1974 through the levy cross sections. • 25% of PG&E’s daily peak winter demand is supplied by Mc Donald Island. The loss of supply from Mc Donald Island would cost between $200 million to $1 billion. McDonald Island levy breech Levee Breech = Scour Scour is bad for buried structures! Levee Break Dimensions Probable Max. Scour Island Date of Flood Cause of Levee Failure Width Scour Depth Length Lower Jones Tract 1980 Levee Failure - Rodents? Webb Tract 1980 High Water Failure 800 ft. 3800 ft. Holland Tract 1980 High Water Failure 300 ft. 2600 ft. McDonald Island August 23, 1982 Levee Failure - Rodents? 650 ft. -70 ft. 1200 ft. Venice Island November 1982 High Water Failure 510 ft. -35 ft. 2200 ft. Mildred Island November 1982 High Water Failure 450 ft. -90 ft. 550 ft. Bradford Island 1983 High Water Failure New Hope Tract February 1986 High Water Failure McCormick-Williams Tract February 1986 Overtopping Deadhorse Island February 1986 Overtopping 190 ft. None None Glanville Tract February 1986 Overtopping None None Little Mandeville Island February 1986 High Water Failure 190 ft. -25 ft. 200 ft. Tyler Island February 21, 1986 Overtopping 375 ft. -45 ft. 1900 ft. Upper Jones Tract June 3, 2004 Levee Failure - Rodents? 260 ft. -50 ft. But How Deep? • Pipe Spec: 24” DSAW, 0.750 WT, Gr. X-70, w/ 16 mils FBE (Mfg: Nippon Steel) • Open Cut Coating: 1” concrete added over FBE for buoyancy control • HDD Coating: 40 mils Powercrete over FBE HDD Design • Pipeline outside of modeled scour zone. -

San Joaquin River Riparian Habitat Below Friant Dam: Preservation and Restoration1
SAN JOAQUIN RIVER RIPARIAN HABITAT BELOW FRIANT DAM: PRESERVATION AND RESTORATION1 Donn Furman2 Abstract: Riparian habitat along California's San Joa- quin River in the 25 miles between Friant Darn and Free- Table 1 – Riparian wildlife/vegetation way 99 occurs on approximately 6 percent of its his- corridor toric range. It is threatened directly and indirectly by Corridor Corridor increased urban encroachment such as residential hous- Category Acres Percent ing, certain recreational uses, sand and gravel extraction, Water 1,088 14.0 aquiculture, and road construction. The San Joaquin Trees 588 7.0 River Committee was formed in 1985 to advocate preser- Shrubs 400 5.0 Other riparianl 1,844 23.0 vation and restoration of riparian habitat. The Com- Sensitive Biotic2 101 1.5 mittee works with local school districts to facilitate use Agriculture 148 2.0 of riverbottom riparian forest areas for outdoor envi- Recreation 309 4.0 ronmental education. We recently formed a land trust Sand and gravel 606 7.5 called the San Joaquin River Parkway and Conservation Riparian buffer 2,846 36.0 Trust to preserve land through acquisition in fee and ne- Total 7,900 100.0 gotiation of conservation easements. Opportunities for 1 Land supporting riparian-type vegetation. In increasing riverbottom riparian habitat are presented by most cases this land has been mined for sand and gravel, and is comprised of lands from which sand and gravel have been extracted. gravel ponds. 2 Range of a Threatened or Endangered plant or animal species. Study Area The majority of the undisturbed riparian habitat lies between Friant Dam and Highway 41 beyond the city limits of Fresno. -

2010-2011 California Regulations for Waterfowl and Upland Game Hunting, Public Lands
Table of Contents CALIFORNIA General Information Contacting DFG ....................................... 2 10-11 Licenses, Stamps, & Permits................... 3 Waterfowl & Upland Shoot Time Tables ................................... 4 Game Hunting and Unlawful Activities .......................... 6 Hunting & Other Public Uses on State & Federal Waterfowl Hunting Lands Regulations Summary of Changes for 10-11 ............... 7 Seasons and Limits ................................. 9 Effective July 1, 2010 - June 30, 2011 Waterfowl Consumption Health except as noted. Warnings ............................................. 12 State of California Special Goose Hunt Area Maps ............ 14 Governor Arnold Schwarzenegger Waterfowl Zone Map ....Inside Back Cover Natural Resources Agency Upland Game Bird, Small Game Secretary Lester A. Snow Mammal, and Crow Hunting Regulation Summary ............................. 16 Fish and Game Commission President Jim Kellogg Seasons & Limits Table ......................... 17 Vice President Richard B. Rogers Hunt Zones ............................................ 18 Commissioner Michael Sutton Hunting and Other Public Uses on Commissioner Daniel W. Richards State and Federal Areas Acting Executive Director Jon Fischer Reservation System .............................. 20 General Public Use Activities on Department of Fish and Game Director John McCamman All State Wildlife Areas ....................... 23 Hunting, Firearms, and Archery Alternate communication formats are available upon request. If reasonable -

GRA 9 – South Delta
2-900 .! 2-905 .! 2-950 .! 2-952 2-908 .! .! 2-910 .! 2-960 .! 2-915 .! 2-963 .! 2-964 2-965 .! .! 2-917 .! 2-970 2-920 ! .! . 2-922 .! 2-924 .! 2-974 .! San Joaquin County 2-980 2-929 .! .! 2-927 .! .! 2-925 2-932 2-940 Contra Costa .! .! County .! 2-930 2-935 .! Alameda 2-934 County ! . Sources: Esri, DeLorme, NAVTEQ, USGS, Intermap, iPC, NRCAN, Esri Japan, METI, Esri China (Hong Kong), Esri (Thailand), TomTom, 2013 Calif. Dept. of Fish and Wildlife Area Map Office of Spill Prevention and Response I Data Source: O SPR NAD_1983_C alifornia_Teale_Albers ACP2 - GRA9 Requestor: ACP Coordinator Author: J. Muskat Date Created: 5/2 Environmental Sensitive Sites Section 9849 – GRA 9 South Delta Table of Contents GRA 9 Map ............................................................................................................................... 1 Table of Contents ...................................................................................................................... 2 Site Index/Response Action ...................................................................................................... 3 Summary of Response Resources for GRA 9......................................................................... 4 9849.1 Environmentally Sensitive Sites 2-900-A Old River Mouth at San Joaquin River....................................................... 1 2-905-A Franks Tract Complex................................................................................... 4 2-908-A Sand Mound Slough .................................................................................. -

Introduction
INTRODUCTION The purpose of this book is twofold: to provide general information for anyone interested in the California islands and to serve as a field guide for visitors to the islands. The book covers both general history and nat- ural history, from the geological origins of the islands through their aboriginal inhabitants and their marine and terrestrial biotas. Detailed coverage of the flora and fauna of one island alone would completely fill a book of this size; hence only the most common, most readily observed, and most interesting species are included. The names used for the plants and animals discussed in this book are the most up-to-date ones available, based on the scientific literature and the most recently published guidebooks. Common names are always subject to local variations, and they change constantly. Where two names are in common use, they are both mentioned the first time the organism is discussed. Ironically, in recent years scientific names have changed more recently than common names, and the reader concerned about a possible discrepancy in nomenclature should consult the scientific literature. If a significant nomenclatural change has escaped our notice, we apologize. For plants, our primary reference has been The Jepson Manual: Higher Plants of California, edited by James C. Hickman, including the latest lists of errata. Variation from the nomenclature in that volume is due to more recent interpretations, as explained in the text. Certain abbreviations used throughout the text may not be immedi- ately familiar to the general reader; they are as follows: sp., species (sin- gular); spp., species (plural); n. -

Transitions for the Delta Economy
Transitions for the Delta Economy January 2012 Josué Medellín-Azuara, Ellen Hanak, Richard Howitt, and Jay Lund with research support from Molly Ferrell, Katherine Kramer, Michelle Lent, Davin Reed, and Elizabeth Stryjewski Supported with funding from the Watershed Sciences Center, University of California, Davis Summary The Sacramento-San Joaquin Delta consists of some 737,000 acres of low-lying lands and channels at the confluence of the Sacramento and San Joaquin Rivers (Figure S1). This region lies at the very heart of California’s water policy debates, transporting vast flows of water from northern and eastern California to farming and population centers in the western and southern parts of the state. This critical water supply system is threatened by the likelihood that a large earthquake or other natural disaster could inflict catastrophic damage on its fragile levees, sending salt water toward the pumps at its southern edge. In another area of concern, water exports are currently under restriction while regulators and the courts seek to improve conditions for imperiled native fish. Leading policy proposals to address these issues include improvements in land and water management to benefit native species, and the development of a “dual conveyance” system for water exports, in which a new seismically resistant canal or tunnel would convey a portion of water supplies under or around the Delta instead of through the Delta’s channels. This focus on the Delta has caused considerable concern within the Delta itself, where residents and local governments have worried that changes in water supply and environmental management could harm the region’s economy and residents. -

Workshop Report—Earthquakes and High Water As Levee Hazards in the Sacramento-San Joaquin Delta
Workshop report—Earthquakes and High Water as Levee Hazards in the Sacramento-San Joaquin Delta Delta Independent Science Board September 30, 2016 Summary ......................................................................................................................................... 1 Introduction ..................................................................................................................................... 1 Workshop ........................................................................................................................................ 1 Scope ........................................................................................................................................... 1 Structure ...................................................................................................................................... 2 Participants and affiliations ........................................................................................................ 2 Highlights .................................................................................................................................... 3 Earthquakes ............................................................................................................................. 3 High water ............................................................................................................................... 4 Perspectives.................................................................................................................................... -

Comparing Futures for the Sacramento-San Joaquin Delta
comparing futures for the sacramento–san joaquin delta jay lund | ellen hanak | william fleenor william bennett | richard howitt jeffrey mount | peter moyle 2008 Public Policy Institute of California Supported with funding from Stephen D. Bechtel Jr. and the David and Lucile Packard Foundation ISBN: 978-1-58213-130-6 Copyright © 2008 by Public Policy Institute of California All rights reserved San Francisco, CA Short sections of text, not to exceed three paragraphs, may be quoted without written permission provided that full attribution is given to the source and the above copyright notice is included. PPIC does not take or support positions on any ballot measure or on any local, state, or federal legislation, nor does it endorse, support, or oppose any political parties or candidates for public office. Research publications reflect the views of the authors and do not necessarily reflect the views of the staff, officers, or Board of Directors of the Public Policy Institute of California. Summary “Once a landscape has been established, its origins are repressed from memory. It takes on the appearance of an ‘object’ which has been there, outside us, from the start.” Karatani Kojin (1993), Origins of Japanese Literature The Sacramento–San Joaquin Delta is the hub of California’s water supply system and the home of numerous native fish species, five of which already are listed as threatened or endangered. The recent rapid decline of populations of many of these fish species has been followed by court rulings restricting water exports from the Delta, focusing public and political attention on one of California’s most important and iconic water controversies.