BMC Genetics Biomed Central
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) 30.03.2009 /EN FCI-Standard N° 93 PORTUGUESE SHEEPDOG (Cão Da Serra De Aires)
FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B – 6530 Thuin (Belgique) ______________________________________________________________________________ 30.03.2009 /EN FCI-Standard N° 93 PORTUGUESE SHEEPDOG (Cão da Serra de Aires) This illustration does not necessarily show the ideal example of the breed. 2 TRANSLATION : Portuguese Kennel Club. Revised by Raymond Triquet, Jennifer Mulholland and Renée Sporre-Willes. ORIGIN : Portugal. DATE OF PUBLICATION OF THE ORIGINAL VALID STANDARD : 04.11.2008. UTILIZATION : Sheepdog for herding and watching over livestock. FCI-CLASSIFICATION : Group 1 Sheepdogs and Cattle Dogs (except Swiss Cattle Dogs). Section 1 Sheepdogs. Without working trial. BRIEF HISTORICAL SUMMARY : Sheepdog used in the Alentejo region for herding and watching different kinds of livestock; sheep, cattle, horses, goats and pigs. An austere and rustic dog perfectly adapted to the area’s temperature changes and with great endurance for covering long distances herding livestock through the Alentejo plains. GENERAL APPEARANCE : Medium sized dog, medium long (Sub-longilinear), with appreciable rusticity and sobriety, extremely agile and swift, with ample and suspended movement. Long hair of goat-like texture, without undercoat. It has simian-like attitudes and appearance, which is why it is known in its native region as “monkey dog”. IMPORTANT PROPORTIONS : The length of the body is approximately 10% more than the height at the withers. St-FCI n°93/30.03.2009 3 The depth of chest is less than 50% of height at the withers. The muzzle length is 2/3 of the skull length; the width of the skull is slightly less than its length. BEHAVIOUR/TEMPERAMENT : Exceptionally intelligent and very lively. -

IN RAKVERE 17Th & 18Th of October 2020
TWO NATIONAL ALL BREEDS DRIVE IN DOG SHOWS th th IN RAKVERE 17 & 18 of October 2020 Indoor Show will take place in RAKVERE SPORTSHALL, KASTANI 12 RAKVERE, ESTONIA Judging starts at 10 These double national shows are held under the protection of the Estonian Kennel Union. Dogs in intermediate, open, working and champion class compete for the Estonian CAC. Certificate (CAC) winning dog becomes an Estonian champion if the dog is a champion in its own country. Dogs in junior class (9-18 month) compete for the Estonian Junior Certificate – EST JUN CAC and for the Estonian Junior Champion title. Dogs in veteran class compete for the Estonian veteran certificate – EST VET CAC and for the Estonian Veteran Champion title. Organizer: RAKVERE KOERTESÕPRADE KLUBI INFO: by phone +372 521 9294 Maire Tõnurist CLASSES: /The decisive date concerning the age is the day of the show/ Babypuppy (4-6 month) BOB BABY BABY BIS Puppy (6-9 month) BOB PUPPY PUPPY BIS All dogs (except baby and puppy) Junior (9-18 month) compete for JUN-CAC BOB JUN JUN BIS in junior, intermediate, Intermediate (15-24 month) open, working, champion Open (from 15 month) Compete and veteran class Working (from 15 month) enclose the working certificate for CAC can compete for BOB, BIG Champion (from 15 month) enclose the champion title Veteran (from 8 year) VET CACEST VET CH title + BOB VET VET BIS and Best In Show ! ADDITIONAL COMPETITIONS: All dogs in competitions must be at least 9 month old and take part from this show and get also at least „very good“ from breed judging to take part at these additional competitions (except breeding dog himself in progeny class) Brace competition - for a male and a female dog of the same breed owned by the same owner. -

AKC Education Webinar Series: FSS/Miscellaneous Questions and Answers
AKC Education Webinar Series: FSS/Miscellaneous Questions and Answers 1. Can an FSS Parent Club offer an ATT Test? If the Parent Club is licensed to hold any event, it may offer an ATT Test. 2. For a domestically evolving breed that started with an independent registry for 20 years, then accepted into UKC for an additional 20 years, can these two be combined to qualify for the 40 years required to be considered into FSS, or must the 40 be done within the same registry body? The history of a breed having a registry for a minimum of 40 years can be merged as described, staff will review to determine if it would meet the parameters required. 3. Does AKC have reciprocity with UKC? AKC has open registration with individual breeds with that are registered by UKC. A breed requesting to be enrolled in the AKC FSS based upon UKC recognition would have reciprocity, if it meets the number of years being in existence, until the breed becomes AKC recognized. At which time the individual breeds may request to keep the studbook open for UKC registered dogs. 4. Is it a good practice to submit all of the required data to AKC in one PDF electronically to FSS as a final submission, once all criteria is met for the ease of processing? The FSS Department keeps track of required data, meeting minutes maybe a PDF, the other requirements need to be in different formats. Breed Standard needs to be a word document, membership list needs to be in an Excel as provided. -

FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) 30.03.2009 /EN FCI-Standard N° 93 CÃO DA SERRA DE AIRES (Portuguese Sheepdog)
FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1er B – 6530 Thuin (Belgique) ______________________________________________________________________________ _______________________________________________________________ 30.03.2009 /EN _______________________________________________________________ FCI-Standard N° 93 CÃO DA SERRA DE AIRES (Portuguese Sheepdog) . 2 TRANSLATION: Portuguese Kennel Club. Revised by Raymond Triquet, Jennifer Mulholland and Renée Sporre-Willes. Official language (EN). ORIGIN: Portugal. DATE OF PUBLICATION OF THE ORIGINAL VALID STANDARD: 04.11.2008. UTILIZATION: Sheepdog for herding and watching over livestock. FCI-CLASSIFICATION: Group 1 Sheepdogs and Cattle Dogs (except Swiss Cattle Dogs). Section 1 Sheepdogs. Without working trial. BRIEF HISTORICAL SUMMARY: Sheepdog used in the Alentejo region for herding and watching different kinds of livestock; sheep, cattle, horses, goats and pigs. An austere and rustic dog perfectly adapted to the area’s temperature changes and with great endurance for covering long distances herding livestock through the Alentejo plains. GENERAL APPEARANCE: Medium sized dog, medium long (Sub-longilinear), with appreciable rusticity and sobriety, extremely agile and swift, with ample and suspended movement. Long hair of goat-like texture, without undercoat. It has simian-like attitudes and appearance, which is why it is known in its native region as “monkey dog”. IMPORTANT PROPORTIONS: The length of the body is approximately 10% more than the height at the withers. St-FCI n°93/30.03.2009 3 The depth of chest is less than 50% of height at the withers. The muzzle length is 2/3 of the skull length; the width of the skull is slightly less than its length. BEHAVIOUR/TEMPERAMENT: Exceptionally intelligent and very lively. Very devoted to the shepherd and the herd, it can be somewhat wary of strangers and vigilant at night. -

(PT) & Herding Instinct Tests SUNDAY, Aug 29, 2021 AKC Farm D
NOTES: FDC AND HERDING TESTS PREMIUM The picture can't be displayed. When not competing, dogs must be confined or held on leash and kept away from the Entries close at Trial Secretary’s Office Wednesday, August 18, 2021 at livestock. Under no circumstances will dogs be allowed to disturb the resting stock or 6:00 pm CST, or when numerical limit per judge or stock limitations are interfere outside the ring with another dog’s performance in the ring. reached. NOTE: Entries for FDC and Herding Instinct will be accepted on the day of the tests space permitting. HT and PT are pre-registration only. No unentered dogs (except set-out dogs) will be allowed at the event site. Drawing for running order will be held on August 18, 2021 at 6:00 p.m. at the Trial secretary’s office at 6008 Maurys Trail, Austin, TX 78730 No Alcoholic beverages allowed. The Texas Herding Association and the owners of Destiny Farm assume no responsibility for any loss, damage or injury sustained by exhibitors, handlers or spectators, or by any of their dogs or property. BRING YOUR OWN SHADE TENTS AND CHAIRS, UMBRELLAS AND RAIN GEAR. EMERGENCY PROCEDURES In case of inclement weather or tornado or lightning, exhibitors are to go either in their Licensed by the American Kennel Club cars or head to the barn with their dogs where they can be crated inside the barn. The barn can withstand over 100 mile per hour winds and is built to stay standing. SATURDAY, Aug 28, 2021 Evacuation route is through the front gate turning left to head to Bertram or Burnet or Austin. -

Contact Details Number of Breeds Within FCI Groups List of Breeds
Contact details Last and First name of Judge Szczepańska-Korpetta Joanna Address 08-443 Sobienie Jeziory, Sobienie Kiełczewskie II - 17, Poland E-mail [email protected] Mobile (+48) 600 451 217 Home phone, Fax (+48) 22 839 65 84 ZKwP Branch Warszawa Languages Polish, German Number of breeds within FCI Groups FCI Group FCI group name Level 1 Sheepdogs and Cattle Dogs (except Swiss Cattle Dogs) (1/52) National 2 Pinscher and Schnauzer - Molossoid breeds - Swiss Mountain and Cattle Dogs International and other breeds (54/54) 3 Terriers (34/34) International 5 Spitz and primitive types (61/61) International 8 Retrievers - Flushing Dogs - Water Dogs (22/22) International 9 Companion and Toy Dogs (33/33) International List of breeds FCI Group Breed Level Sheepdogs and Cattle Dogs (except Swiss Cattle Dogs) 1 Polish Lowland Sheepdog (FCI - 251) National Pinscher and Schnauzer - Molossoid breeds - Swiss Mountain and Cattle Dogs and other breeds 2 Affenpinscher (FCI - 186) International 2 Alentejo Mastiff (FCI - 96) International 2 Appenzell Cattle Dog (FCI - 46) International 2 Atlas Mountain Dog - Aidi (FCI - 247) International 2 Austrian Pinscher (FCI - 64) International 2 Bernese Mountain Dog (FCI - 45) International 2 Bosnian and Herzegovinian - Croatian Shepherd Dog (FCI - 355) International 2 Boxer (FCI - 144) International 2 Broholmer (FCI - 315) International 2 Bulldog (FCI - 149) International 2 Bullmastiff (FCI - 157) International 2 Castro Laboreiro Dog (FCI - 170) International 2 Caucasian Shepherd Dog (FCI - 328) International 1 FCI Group -

Qualified for Crufts 2019
Qualified for Crufts 2019 at the Benelux Winner Show in Amsterdam 9th August 2018 Breed DogName ChipTattoo Gender Owner Australian Kelpie Yaparoos Arramagong Taz 528140000622424 NHSB 3013958 M Marielle Van Der Heeden Australian Kelpie Didaktic's Omg 978101081715689 FI 39682/17 F Marika Lehtonen Belgian Shepherd Dog, Groenendael Revloch Figo 956000008566119 IKC Z48074 M Jean Lawless Belgian Shepherd Dog, Groenendael Revloch Karma Of Woofhouse 941000017117451 IKC Z84192 F Pamela Mckenzie-Hewitt Belgian Shepherd Dog, Laekenois Ruby Rivers Tristan 968000005743426 VDH SE53174/2011 M Karl Strugalla Belgian Shepherd Dog, Malinois Alcosetha'S Jalco 528140000572448 SE 46836/2014 M Ylva Persson Belgian Shepherd Dog, Malinois Boholdt Gloria Gaynor 208250000102703 DKK 18423/2017 F Annelise Sindalsen Belgian Shepherd Dog, Tervueren Glimbi S El Ninõ V. Ginger 208210000413061 DKK DK07707/2013 M Erik Breukelman Belgian Shepherd Dog, Tervueren Ketunpolun Kiss Me Kate 978101081395539 LSVK BST0051/15 F Marciukaitiene Asta Schipperke Slavjan Al Capone 643094100436648 RKF 3977884 M Maria Spiridonova Schipperke BEACHVIEW'S TIL DEATH DO US PART 1653392 AKC NP33073101 F Lindsey @ Joanne Mclachlan Czechoslovakian Wolfdog My Kayden De La Legende Du Loup Noir 250268500976212 LOF 4004/0 M Regis Haydo Czechoslovakian Wolfdog Arimminum Yasmeen 380260010336510 ROI 15/155486 F Scaglia Laura German Shepherd Dog, double coat Omero Haus Dando 804098100057016 UKU 0182383 M Kholodna Ievgeniia German Shepherd Dog, double coat Adele Da Soc Port 620098100741122 LOP 507637 F -

Canis-Adn-Mix Razas Caninas
CANIS-ADN-MIX RAZAS CANINAS Affenpinscher Bohemian Shepherd Afghan Hound Bolognese Airedale Terrier Border Collie Akita Border Terrier Alaskan Klee Kai Borzoi Alaskan Malamute Boston Terrier Alpine Dachsbracke Bouvier Des Flanders American English Coonhound Boxer American Eskimo Dog Boykin Spaniel American Foxhound Bracco Italiano American Hairless Rat Terrier Braque du Bourbonnais American Indian Dog Brazilian Terrier American Staffordshire Terrier Briard American Water Spaniel Brussels Griffon Anatolian Shepherd Dog Bulgarian Shepherd Appenzell Cattle Dog Bull Arab Argentine Dogo Bull Terrier (Miniature) Australian Cattle Dog Bull Terrier (Standard) Australian Kelpie Bulldog (American) Australian Labradoodle Bulldog (Standard) Australian Shepherd Bullmastiff Australian Stumpy Tail Cattle Dog Cairn Terrier Australian Terrier Canaan Dog Azawakh Canadian Eskimo Dog Barbet Cane Corso Basenji Cardigan Welsh Corgi Basset Bleu de Gascogne Catahoula Leopard Dog Basset Fauve de Bretagne Catalan Sheepdog Basset Griffon Vendeen (Grand) Caucasian Shepherd Dog Basset Griffon Vendeen (Petit) Cavalier King Charles Spaniel Basset Hound Central Asian Ovcharka Bavarian Mountain Hound Cesky Terrier Beagle Chesapeake Bay Retriever Bearded Collie Chihuahua Beauceron Chinese Crested Bedlington Terrier Chinese Shar-Pei Belgian Malinois Chinook Belgian Mastiff Chow Chow Belgian Sheepdog Cirneco del Etna Belgian Tervuren Clumber Spaniel Bergamasco Cocker Spaniel Berger Picard Collie Bernese Mountain Dog Continental Toy Spaniel (phalene and papillon) Bichon -
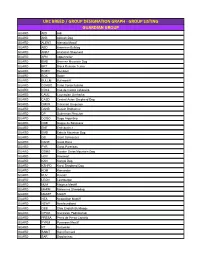
Ukc Breed / Group Designation Graph
UKC BREED / GROUP DESIGNATION GRAPH - GROUP LISTING GUARDIAN GROUP GUARD AIDI Aidi GUARD AKB Akbash Dog GUARD ALENT Alentejo Mastiff GUARD ABD American Bulldog GUARD ANAT Anatolian Shepherd GUARD APN Appenzeller GUARD BMD Bernese Mountain Dog GUARD BRT Black Russian Terrier GUARD BOER Boerboel GUARD BOX Boxer GUARD BULLM Bullmastiff GUARD CORSO Cane Corso Italiano GUARD CDCL Cao de Castro Laboreiro GUARD CAUC Caucasian Ovcharka GUARD CASD Central Asian Shepherd Dog GUARD CMUR Cimarron Uruguayo GUARD DANB Danish Broholmer GUARD DP Doberman Pinscher GUARD DOGO Dogo Argentino GUARD DDB Dogue de Bordeaux GUARD ENT Entlebucher GUARD EMD Estrela Mountain Dog GUARD GS Giant Schnauzer GUARD DANE Great Dane GUARD PYR Great Pyrenees GUARD GSMD Greater Swiss Mountain Dog GUARD HOV Hovawart GUARD KAN Kangal Dog GUARD KSHPD Karst Shepherd Dog GUARD KOM Komondor GUARD KUV Kuvasz GUARD LEON Leonberger GUARD MJM Majorca Mastiff GUARD MARM Maremma Sheepdog GUARD MASTF Mastiff GUARD NEA Neapolitan Mastiff GUARD NEWF Newfoundland GUARD OEB Olde English Bulldogge GUARD OPOD Owczarek Podhalanski GUARD PRESA Perro de Presa Canario GUARD PYRM Pyrenean Mastiff GUARD RT Rottweiler GUARD SAINT Saint Bernard GUARD SAR Sarplaninac GUARD SC Slovak Cuvac GUARD SMAST Spanish Mastiff GUARD SSCH Standard Schnauzer GUARD TM Tibetan Mastiff GUARD TJAK Tornjak GUARD TOSA Tosa Ken SCENTHOUND GROUP SCENT AD Alpine Dachsbracke SCENT B&T American Black & Tan Coonhound SCENT AF American Foxhound SCENT ALH American Leopard Hound SCENT AFVP Anglo-Francais de Petite Venerie SCENT -

American Pit Bull Terrier
Portuguese Sheepdog Official UKC Breed Standard Herding Dog Group ©Copyright 2006, United Kennel Club CHARACTERISTICS Very lively and intelligent, devoted to the flock and the shepherd but wary of strangers and a good night guarding dog. HEAD Strong and broad, not overly long, and never round. The stop is well-defined, and the toplines of the skull and muzzle are divergent. SKULL - Slightly longer than wide, convex on the sides, but flat between the ears. There is a pronounced median furrow that runs halfway up the forehead. The occiput is prominent. MUZZLE - Short, only about two-thirds the length of The goals and purposes of this breed standard the skull. The nasal bone is straight or slightly include: to furnish guidelines for breeders who wish hollowed. The lips are tight and thin and do not to maintain the quality of their breed and to improve overlap. it; to advance this breed to a state of similarity TEETH – The Portuguese Sheepdog has a complete set throughout the world; and to act as a guide for of evenly spaced, white teeth meeting in a scissors judges. bite. Breeders and judges have the responsibility to Disqualifications: Undershot or overshot. avoid any conditions or exaggerations that are NOSE - Slightly raised from the profile of the foreface, detrimental to the health, welfare, essence and with large nostrils. Black color is preferred, but others soundness of this breed, and must take the are acceptable as long as it is darker than the color of responsibility to see that these are not perpetuated. the coat. Any departure from the following should be EYES - Medium size and rounded in shape. -

Contact Details Number of Breeds Within FCI Groups List of Breeds
Contact details Last and First name of Judge Kot Andrzej Address 42-600 Tarnowskie Góry, Torowa 58, Poland E-mail [email protected] Mobile (+48) 601 428 154 Home phone, Fax ZKwP Branch Bytom Languages Polish Number of breeds within FCI Groups FCI Group FCI group name Level 1 Sheepdogs and Cattle Dogs (except Swiss Cattle Dogs) (2/52) International 2 Pinscher and Schnauzer - Molossoid breeds - Swiss Mountain and Cattle Dogs International and other breeds (41/54) 6 Scenthounds and related breeds (2/77) International 10 Sighthounds (1/13) International List of breeds FCI Group Breed Level Sheepdogs and Cattle Dogs (except Swiss Cattle Dogs) 1 Polish Lowland Sheepdog (FCI - 251) International 1 Tatra Shepherd Dog (FCI - 252) International Pinscher and Schnauzer - Molossoid breeds - Swiss Mountain and Cattle Dogs and other breeds 2 Alentejo Mastiff (FCI - 96) International 2 Appenzell Cattle Dog (FCI - 46) International 2 Atlas Mountain Dog - Aidi (FCI - 247) International 2 Bernese Mountain Dog (FCI - 45) International 2 Bosnian and Herzegovinian - Croatian Shepherd Dog (FCI - 355) International 2 Broholmer (FCI - 315) International 2 Bulldog (FCI - 149) International 2 Bullmastiff (FCI - 157) International 2 Castro Laboreiro Dog (FCI - 170) International 2 Caucasian Shepherd Dog (FCI - 328) International 2 Central Asia Shepherd Dog (FCI - 335) International 2 Ciobanesc Romanesc de Bucovina (FCI - 357) International 2 Dogo Argentino (FCI - 292) International 2 Dogue de Bordeaux (FCI - 116) International 1 FCI Group Breed Level 2 Entlebuch -

AUSTRALIAN NATIONAL KENNEL COUNCIL LTD Judge
AUSTRALIAN NATIONAL KENNEL COUNCIL LTD GROUP 1 - TOYS Judge: Affenpinscher Bourbonnais Pointing Dog * Dachshund (Min. Long) Australian Silky Terrier Bracco Italiano Dachshund (Rabbit Long Bichon Frise Brittany Haired) * Bolognese * Chesapeake Bay Retriever Dachshund (Smooth) Cavalier King Charles Spaniel Clumber Spaniel Dachshund (Min. Smooth) Chihuahua (Long Coat) Cocker Spaniel Dachshund (Rabbit Smooth Chihuahua (Smooth Coat) Cocker Spaniel (American) Haired) * Chinese Crested Dog Curly Coated Retriever Dachshund (Wire) Continental Toy Spaniel Deutch Stichelhaar * Dachshund (Min. Wire) (Papillon) Drentsche Partridge Dog * Dachshund (Rabbit Wire Continental Toy Spaniel English Setter Haired) * (Phalene) * English Springer Spaniel Deerhound Coton De Tulear Field Spaniel Drever * Griffon Belge * Flat Coated Retriever Fawn Brittany Griffon * English Toy Terrier (Black & French Pointing Dog Gascogne Finnish Hound * Tan) Type * Finnish Spitz Griffon Bruxellois French Pointing Dog Pyrenean Foxhound Havanese Type * Gascon Saintongeois * Italian Greyhound French Spaniel * German Hound * Japanese Chin French Water Dog * Grand Basset Griffon King Charles Spaniel Frisian Pointing Dog * Vendeen Kromforhlander * Frisian Water Dog * Grand Griffon Vendeen * Lowchen German Shorthaired Pointer Great Anglo French-White & Maltese German Wirehaired Pointer Black Hound * Miniature Pinscher German Spaniel * Great Anglo-French Tri Colour Hound * Pekingese Golden Retriever Great Anglo-French White & Petit Brabancon * Gordon Setter Orange Hound * Pomeranian