Faunal Assemblages on the Pacific-Antarctic Ridge Near The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

4. Deep-Tow Observations at the East Pacific Rise, 8°45N, and Some Interpretations
4. DEEP-TOW OBSERVATIONS AT THE EAST PACIFIC RISE, 8°45N, AND SOME INTERPRETATIONS Peter Lonsdale and F. N. Spiess, University of California, San Diego, Marine Physical Laboratory, Scripps Institution of Oceanography, La Jolla, California ABSTRACT A near-bottom survey of a 24-km length of the East Pacific Rise (EPR) crest near the Leg 54 drill sites has established that the axial ridge is a 12- to 15-km-wide lava plateau, bounded by steep 300-meter-high slopes that in places are large outward-facing fault scarps. The plateau is bisected asymmetrically by a 1- to 2-km-wide crestal rift zone, with summit grabens, pillow walls, and axial peaks, which is the locus of dike injection and fissure eruption. About 900 sets of bottom photos of this rift zone and adjacent parts of the plateau show that the upper oceanic crust is composed of several dif- ferent types of pillow and sheet lava. Sheet lava is more abundant at this rise crest than on slow-spreading ridges or on some other fast- spreading rises. Beyond 2 km from the axis, most of the plateau has a patchy veneer of sediment, and its surface is increasingly broken by extensional faults and fissures. At the plateau's margins, secondary volcanism builds subcircular peaks and partly buries the fault scarps formed on the plateau and at its boundaries. Another deep-tow survey of a patch of young abyssal hills 20 to 30 km east of the spreading axis mapped a highly lineated terrain of inactive horsts and grabens. They were created by extension on inward- and outward- facing normal faults, in a zone 12 to 20 km from the axis. -

Presentation on Pacific Plate and Associated Boundaries
PACIFIC PLATE AND ASSOCIATED BOUNDARIES The Pacific Plate • Pacific Plate is the largest plate and an oceanic plate. • It shares its boundaries with numerous plates namely; North American Plate.(Convergent and transform fault) Philippine Plate.(Convergent) Juan de Fuca Plate.(Convergent) Indo – Australian Plate.(Convergent, Transform Fault) Cocos Plate.(Divergent) Nazca Plate.(Divergent) Antarctic Plate.(Divergent,Transform Fault) Types of Plate Boundaries • Convergent Boundary: Subduction zones where two plates converges. Eg; Aleutian Islands(Alaska) • Divergent Boundary: Spreading centres where two plates move away from each other. Eg; East Pacific Rise (MOR, Pacific Ocean). • Transform Faults: Boundary where two plates slide past each other. For Eg. ; San Andreas Fault. BOUNDARY WITH ANTARCTIC PLATE DIVERGENT BOUNDARY • Pacific – Antarctic Ridge TRANSFORM FAULT • Louisville Seamount Chain Pacific – Antarctic Ridge Pacific – Antarctic Ridge(PAR) is located on the seafloor of the South Pacific Ocean. It is driven by the interaction of a mid oceanic ridge and deep mantle plumes located in the eastern portion of East Pacific Ridge. Louisville Seamount Chain It is the longest line of seamount chain in the Pacific Ocean of about 4,300 km, formed along the transform boundary in the western side between Pacific plate and Antarctic plate. It was formed from the Pacific Plate sliding over a long – lived centre of upwelling magma called the Louisville hotspot. BOUNDARY WITH PHILIPPINE PLATE CONVERGENT BOUNDARY • Izu – Ogasawara Trench • Mariana Trench Izu – Ogasawara Trench It is an oceanic trench in the western Pacific Ocean. It stretches from Japan to northern most section of Mariana Trench. Here, the Pacific Plate is being subducted beneath the Philippine Sea Plate. -

'Sounding the Pacific Ocean; an Echo Sounder Traverse of the Eastern Pacific'
Earthlearningidea - https://www.earthlearningidea.com/ Sounding the Pacific Ocean An echo sounder traverse of the eastern Pacific Ask them to describe out loud the main changes in depth to their partner in the class, and to discuss if they found any of these surprising, and if so, why. (Answers will vary, but the rapid descent into the Peru/Chile Trench may be a surprise: also the monotonously flat section and the sudden rise to the seamount). Ask them to label the following on their graphs: Continental shelf – depth less than 150m Continental slope – depths from 150m to 2000m Peru/Chile Trench – depth of 8000m The harbour at Callao, Lima, Peru in 2005. Abyssal plain – very level sea bed at around A research ship sets out from the coast of Peru, 5000m depth near Lima, and travels due west for 3600km. As Seamount – isolated underwater peak rising to the ship moves, its echo sounder is continuously 2000m or less below sea level recording the depth of the ocean floor below sea East Pacific Rise –an underwater mountain level. Some of the echo sounder readings are range at around 2000m depth shown in the Table. The Table shows the distance Rift valley of the East Pacific Rise – around from Lima in kilometres and the time taken for the 3500m depth. echo sounder’s ‘ping’ to travel from the transmitter Ask what they would expect to happen to the in the ship’s hull and back up again to the water depth as the ship sails a few hundred receiver. We know that the speed of sound in sea kilometres further west. -

High-Resolution Bathymetry of the East Pacific Rise Axial Summit Trough 9O 49’ - 51’N: a Compilation of Alvin Scanning Sonar and Altimetry Data from 1991-1995
1 High-resolution bathymetry of the East Pacific Rise axial summit trough 9o 49’ - 51’N: A compilation of Alvin scanning sonar and altimetry data from 1991-1995. Kurras, G.J., Edwards, M.H., Fornari, D.J. School of Ocean Earth Science and Technology, University of Hawai’i, Honolulu, HI multibeam surveys. At fast-spreading MORs, lobate lava Abstract. Bathymetric data acquired during six cruises to the o flows are generally <<10 m thick and topographic variability East Pacific Rise crest 9 49’-51’N are compiled into a single of constructional volcanic terrain at the axis is subtle. bathymetric database and used to create high-resolution Mapping seafloor structure and monitoring topographic bathymetry maps of two geologically and biologically active changes with repeat surveys on fast spreading MORs thus areas. Scanning sonar and Alvin altimetry were collected requires mapping techniques that can resolve features <<10 m during 23 Alvin dives to the P-vent/Bio9 and Biomarker-141 high and 1 x 102 m2 in area. areas during 1991-95. Bathymetry data were filtered, edited, This paper discusses the compilation, processing, and and processed as individual dives then combined into a single analysis of bathymetry acquired using the submersible Alvin database for each cruise and statistically evaluated. Cruise for two well-studied areas of the EPR crest within the axial data sets were numerically and visually compared and cross- summit collapse trough (ASCT) [Fornari et al., in press], in a correlated to known bottom features and fixed seafloor region which experienced a submarine eruption in 1991 reference markers, then horizontally shifted to establish a [Haymon et al., 1993; Gregg et al., 1996a]. -

SUBDUCTION ZONES • Most Subduction Zones Are Found in the Pacific Ocean
ANOTHER MEXICAN EARTHQUAKE! Magnitude 7.1, Tuesday Sept. 19, 2017 Why is there no oceanic crust older than 200 million years? SUBDUCTION • If new oceanic crust is being continuously created along the earth’s spreading ridge system, then we must find some way to re-cycle it back into the mantle. [WHY? –otherwise the earth would be expanding!!!!] • Old oceanic crust (>200 million years) is returned to the mantle at the deep ocean trenches. • These are known as SUBDUCTION ZONES • Most subduction zones are found in the Pacific Ocean. This means that the Pacific Ocean is shrinking and the Atlantic Ocean is expanding. Convection in the mantle Lithosphere Recap Lithosphere (or plate) – is rigid and is composed of crust and upper mantle. Thickness varies from 10-150 km. Asthenosphere – is soft, plastic and convecting. Melting of the asthenosphere produces volcanic rocks at ocean ridges. SUBDUCTION Cross-section through the southern Pacific Ocean New oceanic crust and lithosphere are created at the East Pacific Rise Old oceanic crust and lithosphere are subducted at deep ocean trenches (Tonga trench and Chile trench). Kamchatka Pacific Ocean trench Aleutian trench Japan trench Costa Rica trench Marianas trench Tonga trench Chile trench East Pacific Rise Notice that the trenches are often curved with the convex side facing the direction of subduction. This is because the earth is spherical Consequently by observing the curvature of the trench we can easily determine which side is being subducted Details of a subduction zone • Slab of lithosphere descends back into the mantle at a deep ocean trench. • Earthquakes trace the descent of the slab into the mantle (Benioff Zone). -

Late Cretaceous and Paleogene Tectonic Evolution of the North Pacific Ocean
Earth and Planetary Science Letters, 65 (1983) 145-166 145 Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands [4l Late Cretaceous and Paleogene tectonic evolution of the North Pacific Ocean David K. Rea l and John M. Dixon 2 I Oceanography Program, Department of Atmospheric and Oceanic Science, University of Michigan, Ann Arbor, MI 48109 (U.S.A.) 2 Department of Geological Sciences, Queen's University, Kingston, Ont. KTL 3N6 (Canada) Received March 24, 1983 Revised version received July 11, 1983 The Late Cretaceous history of the northern Pacific Ocean has not been adequately deciphered, largely because a major plate reorganization occurred during the Cretaceous magnetic quiet interval. Using primary data to reconstruct plate motions from fracture zone trends and Late Cretaceous seafloor spreading magnetic anomalies allows formulation of a reasonable sequence of events that accounts for all the geologic features of that region, especially the Emperor and Chinook troughs. The primary event in our reconstruction is the subduction of the old northwest Pacific triple junction. New relative plate motions imposed by formation of convergent boundaries along both the northern Pacific and Farallon plates caused the Farallon plate to crack. This subdivision occurred 82 m.y. ago and resulted in the formation of the Kula and Chinook plates. The Chinook plate was bounded on the north by the Chinook-Kula ridge, the western arm of the Great Magnetic Bight, on the west by the southern Emperor trough, a slowly spreading rift valley, on the south by the Mendocino transform, and on the east by the Chinook-Farallon ridge. -

Drilling the Crust at Mid-Ocean Ridges
or collective redistirbution of any portion of this article by photocopy machine, reposting, or other means is permitted only with the approval of Th e Oceanography Society. Send all correspondence to: [email protected] or Th eOceanography PO Box 1931, Rockville, USA.Society, MD 20849-1931, or e to: [email protected] Oceanography correspondence all Society. Send of Th approval portionthe ofwith any articlepermitted only photocopy by is of machine, reposting, this means or collective or other redistirbution articleis has been published in Th SPECIAL ISSUE FEATURE Oceanography Drilling the Crust Threproduction, Republication, systemmatic research. for this and teaching article copy to use in reserved. e is rights granted All OceanographyPermission Society. by 2007 eCopyright Oceanography Society. journal of Th 20, Number 1, a quarterly , Volume at Mid-Ocean Ridges An “in Depth” perspective BY BENOÎT ILDEFONSE, PETER A. RONA, AND DONNA BLACKMAN In April 1961, 13.5 m of basalts were drilled off Guadalupe In the early 1970s, almost 15 years after the fi rst Mohole Island about 240 km west of Mexico’s Baja California, to- attempt, attendees of a Penrose fi eld conference (Confer- gether with a few hundred meters of Miocene sediments, in ence Participants, 1972) formulated the concept of a layered about 3500 m of water. This fi rst-time exploit, reported by oceanic crust composed of lavas, underlain by sheeted dikes, John Steinbeck for Life magazine, aimed to be the test phase then gabbros (corresponding to the seismic layers 2A, 2B, for the considerably more ambitious Mohole project, whose and 3, respectively), which themselves overlay mantle perido- objective was to drill through the oceanic crust down to tites. -

A Plate Model for Jurassic to Recent Intraplate Volcanism in the Pacific Ocean Basin
A Plate Model for Jurassic to Recent Intraplate Volcanism in the Pacific Ocean Basin Alan D. Smith Department of Geological Sciences, University of Durham, Durham, DH1 3LE, UK Email: [email protected] 1 ABSTRACT Reconstruction of the tectonic evolution of the Pacific basin indicates a direct relationship between intraplate volcanism and plate reorganisations, which suggests volcanism was controlled by fracturing and extension of the lithosphere. Middle Jurassic to Early Cretaceous intraplate volcanism included oceanic plateau formation at triple junctions (Shatsky Rise, western Mid Pacific Mountains) and a diffuse pattern of ocean island volcanism (Marcus Wake, Magellan seamounts) reflecting an absence of any well-defined stress field within the plate. The stress field changed in the Early Cretaceous when accretion of the Insular terrane to the North American Cordillera and the Median Tectonic arc to New Zealand, stalled migration of the Pacific- Farallon and Pacific-Phoenix ocean ridges, leading to the generation of the Ontong Java, Manahiki, Hikurangi and Hess Rise oceanic plateaus. Plate reorganisations in the Late Cretaceous resulted from the breakup of the Phoenix and Izanagi plates through collision of the Pacific-Phoenix ocean ridge with the southwest margin of the basin, and development of island arc-marginal basin systems in the northwest of the basin. The Pacific plate nonetheless remained largely bounded by spreading centres, and intraplate volcanism followed pre-existing lines of weakness in the plate fabric (Line Islands), or resulted from fractures generated by ocean ridge subduction beneath island arc systems (Emperor chain). The Pacific plate began to subduct under Asia in the Early Eocene from the record of accreted material along the Japanese margin. -

Oceanographyra Spocietyhy
OceThe OfficiaaL MaganZineog of the Oceanographyra Spocietyhy CITATION Rubin, K.H., S.A. Soule, W.W. Chadwick Jr., D.J. Fornari, D.A. Clague, R.W. Embley, E.T. Baker, M.R. Perfit, D.W. Caress, and R.P. Dziak. 2012. Volcanic eruptions in the deep sea. Oceanography 25(1):142–157, http://dx.doi.org/10.5670/oceanog.2012.12. DOI http://dx.doi.org/10.5670/oceanog.2012.12 COPYRIGHT This article has been published inOceanography , Volume 25, Number 1, a quarterly journal of The Oceanography Society. Copyright 2012 by The Oceanography Society. All rights reserved. USAGE Permission is granted to copy this article for use in teaching and research. Republication, systematic reproduction, or collective redistribution of any portion of this article by photocopy machine, reposting, or other means is permitted only with the approval of The Oceanography Society. Send all correspondence to: [email protected] or The Oceanography Society, PO Box 1931, Rockville, MD 20849-1931, USA. downLoaded from http://www.tos.org/oceanography OCEANIC SPREADING CENTER PROCESSES | Ridge 2000 PROGRAM RESEARCH Volcanic Eruptions in the Deep Sea BY KEnnETH H. RUBIN, S. ADAM SOULE, WILLIAM W. CHADWICK JR., DANIEL J. FORNARI, DAVID A. CLAGUE, RobERT W. EMBLEY, EDWARD T. BAKER, MICHAEL R. PERFIT, DAVID W. CAREss, AND RobERT P. DZIAK Eruption of molten lapilli, ash, and sulfur-rich fumes at Hades vent, West Mata Volcano. 142 Oceanography | Vol. 25, No. 1 AbsTRACT. Volcanic eruptions are important events in Earth’s cycle of magma eruptions, we also discuss submarine generation and crustal construction. Over durations of hours to years, eruptions eruptions in other settings because produce new deposits of lava and/or fragmentary ejecta, transfer heat and magmatic volcanism is a continuum of conditions volatiles from Earth’s interior to the overlying air or seawater, and significantly and processes across the full range of modify the landscape and perturb local ecosystems. -

1 OCEANOGRAPHY the PACIFIC OCEAN by PROF. A. BALSUBRAMANIAN OUTLINE 1.0 Introduction 1.1 Geographic Setting 1.2 Dimension
OCEANOGRAPHY THE PACIFIC OCEAN BY PROF. A. BALSUBRAMANIAN OUTLINE 1.0 Introduction 1.1 Geographic setting 1.2 Dimension 1.3 Depth 1.4 Principal Arms 1.5 Volume of water 2.0 Historical explorations 3.0 Crustal plates 4.0 Profile of the ocean floor 4.1 Continental shelf 4.2 Continental slope 4.3 Submarine canyons 4.4 Deep ocean floor 4.5 Abyssal plains/hills 4.6 Ring of Fire 4.7 Mid ocean ridges 4.8 Deep ocean trenches 4.9 Seamounts 4.10 Guyots 4.11 Icebergs 4.12 Active volcanoes 4.13 Islands or island arcs 5.0 Sedimentation 6.0 Water masses and Temperature 6.1 Salinity 6.2 Density 7.0 Climate 7.1 Water circulation and Ocean Currents 7.2 Oceanic currents 7.3 Ocean waves 8.0 Ecological zones 8.1 Marine fauna 8.2 Marine flora 8.3 Marine sediments 9.0 Economic mineral resources 9.1 Commerce and shipping 9.2 Ports and terminals 10.0 Marine pollution 10.1 Hazards 1 The Objectives After attending this lesson, the learner should be able to comprehend about the geographic setting of the Pacific ocean, its dimension, associated water masses, morphological features of the ocean floor, very significant conditions of the ocean, sediments, marine life, marine pollution and other hazards. In addition the user should be able to understand, the importance of the Pacific in the context of global activities including the historical oceanographic explorations. 1.0 Introduction The world’s oceanic water masses occupy about 97 per cent of the hydrosphere. -

Going to Extremes
New Zealand American Submarine Ring of Fire 2007 Going to Extremes FOCUS AUDIO/VISUAL MATERIALS Archaea (Optional) Equipment for viewing online or downloaded video of vent communities GRADE LEVEL 9-12 (Biology) TEACHING TIME One or two 45-minute class periods, plus time for FOCUS QUESTION student research What are Archaea, and what is their potential significance in hydrothermal communities of the SEATING ARRANGEMENT Kermadec Arc? Classroom style if students are working individu- ally, or groups of two to four students LEARNING OBJECTIVES Students will be able to define “lipid biomarkers,” MAXIMUM NUMBER OF STUDENTS and explain what the presence of certain bio- 30 markers signifies. KEY WORDS Students will be able to describe Archaea, and Kermadec Arc explain why these organisms are often consid- Hydrothermal field ered to be unusual. Archaea Prokaryote Students will be able to compare and contrast Eukaryote Archaea with bacteria and eukaryotes. Methanogen Methanotroph Students will be able to define methanogen and Chemoautotroph methanotroph, and explain the relevance of these terms to Archaea. BACKGROUND INFORMATION The Submarine Ring of Fire is an arc of active vol- Students will be able to discuss the potential sig- canoes that partially encircles the Pacific Ocean nificance of Archaea in hydrothermal communi- Basin, including the Kermadec and Mariana ties of the Kermadec Arc. Islands in the western Pacific, the Aleutian Islands between the Pacific and Bering Sea, the Cascade MATERIALS Mountains in western North America, and numer- Copies of “Going to Extremes Worksheet,” one ous volcanoes on the western coasts of Central copy for each student or student group America and South America. -
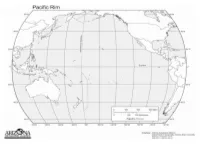
Mapping Ring of Fire Sudent Handout
Mapping the Ring of Fire (part 1) Using a yellow pencil on a map of the world Outline and Label the following in green: make a prediction and shade the areas o United States of America where you think the most earthquakes o Japan occur. Use the information in the data table on the Using an orange pencil on a map of the next page to mark the location of each world make a prediction and shade the earthquake on the world map that follows the areas where you think the most volcanoes data table. occur. Use a blue colored pencil to draw a circle Label each of the following in brown: at each earthquake location. o North America continent o South America continent Use a purple colored pencil to mark the o Australia continent locations of the volcanoes on the map with o Asia continent a triangle. o Pacific Ocean o Sea of Japan o Mt Rainier o Mt Fuji Volcanoes Earthquakes Longitude Latitude Longitude Latitude Longitude Latitude Longitude Latitude 120° W 40° N 125° E 23° N 150° W 60° N 37° E 3° S 110° E 5° S 30° E 35° N 70° W 35° S 145° E 40° N 77° W 4° S 140° E 35° N 120° W 45° N 120° E 10° S 88° E 23° N 12° E 46° N 61° W 15° N 14° E 41° N 121° E 14° S 75° E 28° N 105° W 20° N 105° E 5° S 34° E 7° N 150° W 61° N 75° W 0° 35° E 15° N 74° W 44° N 68° W 47° S 122° W 40° N 70° W 30° S 70° W 30° S 175° E 41° S 30° E 40° N 175° E 39° S 10° E 45° N 121° E 17° N 60° E 30° N 123° E 38° N 85° W 13° N 160° E 55° N Mapping the Ring of Fire (part 2) After you have sketched your inferred View a map of seismic data from the Pacific boundaries compare your map to a map of the Northwest.