Environmental Health Criteria 145 Methyl Parathion
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Additive Interactions of Some Reduced-Risk Biocides and Two
www.nature.com/scientificreports OPEN Additive interactions of some reduced‑risk biocides and two entomopathogenic nematodes suggest implications for integrated control of Spodoptera litura (Lepidoptera: Noctuidae) Rashad Rasool Khan 1,2*, Muhammad Arshad 1*, Asad Aslam 3 & Muhammad Arshad 4 Higher volumes of conventional and novel chemical insecticides are applied by farmers to control resistant strains of armyworm (Spodoperta litura) in Pakistan without knowing their risks to the environment and to public health. Ten reduced‑risk insecticides were tested for their compatibility with two entomopathogenic nematodes (EPNs); Heterorhabditis indica and Steinernema carpocapsae rd against S. litura. The insecticide emamectin benzoate was highly toxic (LC50 = 2.97 mg/l) against 3 instar S. litura larvae when applied alone whereas, novaluron and methoxyfenozide were the least toxic (LC50 = 29.56 mg/l and 21.06 mg/l), respectively. All the insecticides proved harmless against the two EPNs even 96 h after treatment. Indoxacarb, fubendiamide and spinetoram produced the greatest mortalities (72–76%) of S. litura larvae after 72 h when applied in mixtures with H. indica. Lowest mortalities (44.00 ± 3.74% and 48.00 ± 2.89) were observed for mixtures of H. indica with methoxyfenozide and chlorfenapyr, respectively. The positive control treatments with both EPNs (S. carpocapsae and H. indica) produced > 50% mortality 96 h after treatment. For insecticide mixtures with S. carpocapsae, only indoxacarb produced 90% mortality of larvae, whereas, indoxacarb, fubendiamide, emamectin benzoate, and spinetoram produced 90–92% mortality of larvae when applied in mixtures with H. indica. Additive interactions (Chi‑square < 3.84) of EPN mixtures with reduced volumes of reduced‑risk insecticides suggest opportunities to develop more environmentally favorable pest management programs for S. -

Historical Perspectives on Apple Production: Fruit Tree Pest Management, Regulation and New Insecticidal Chemistries
Historical Perspectives on Apple Production: Fruit Tree Pest Management, Regulation and New Insecticidal Chemistries. Peter Jentsch Extension Associate Department of Entomology Cornell University's Hudson Valley Lab 3357 Rt. 9W; PO box 727 Highland, NY 12528 email: [email protected] Phone 845-691-7151 Mobile: 845-417-7465 http://www.nysaes.cornell.edu/ent/faculty/jentsch/ 2 Historical Perspectives on Fruit Production: Fruit Tree Pest Management, Regulation and New Chemistries. by Peter Jentsch I. Historical Use of Pesticides in Apple Production Overview of Apple Production and Pest Management Prior to 1940 Synthetic Pesticide Development and Use II. Influences Changing the Pest Management Profile in Apple Production Chemical Residues in Early Insect Management Historical Chemical Regulation Recent Regulation Developments Changing Pest Management Food Quality Protection Act of 1996 The Science Behind The Methodology Pesticide Revisions – Requirements For New Registrations III. Resistance of Insect Pests to Insecticides Resistance Pest Management Strategies IV. Reduced Risk Chemistries: New Modes of Action and the Insecticide Treadmill Fermentation Microbial Products Bt’s, Abamectins, Spinosads Juvenile Hormone Analogs Formamidines, Juvenile Hormone Analogs And Mimics Insect Growth Regulators Azadirachtin, Thiadiazine Neonicotinyls Major Reduced Risk Materials: Carboxamides, Carboxylic Acid Esters, Granulosis Viruses, Diphenyloxazolines, Insecticidal Soaps, Benzoyl Urea Growth Regulators, Tetronic Acids, Oxadiazenes , Particle Films, Phenoxypyrazoles, Pyridazinones, Spinosads, Tetrazines , Organotins, Quinolines. 3 I Historical Use of Pesticides in Apple Production Overview of Apple Production and Pest Management Prior to 1940 The apple has a rather ominous origin. Its inception is framed in the biblical text regarding the genesis of mankind. The backdrop appears to be the turbulent setting of what many scholars believe to be present day Iraq. -

WO 2010/128115 Al
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization International Bureau (10) International Publication Number (43) International Publication Date 11 November 2010 (11.11.2010) WO 2010/128115 Al (51) International Patent Classification: (81) Designated States (unless otherwise indicated, for every C12N 9/16 (2006.01) kind of national protection available): AE, AG, AL, AM, AO, AT, AU, AZ, BA, BB, BG, BH, BR, BW, BY, BZ, (21) International Application Number: CA, CH, CL, CN, CO, CR, CU, CZ, DE, DK, DM, DO, PCT/EP20 10/056204 DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, (22) International Filing Date: HN, HR, HU, ID, IL, IN, IS, JP, KE, KG, KM, KN, KP, 6 May 2010 (06.05.2010) KR, KZ, LA, LC, LK, LR, LS, LT, LU, LY, MA, MD, ME, MG, MK, MN, MW, MX, MY, MZ, NA, NG, NI, (25) Filing Language: English NO, NZ, OM, PE, PG, PH, PL, PT, RO, RS, RU, SC, SD, (26) Publication Language: English SE, SG, SK, SL, SM, ST, SV, SY, TH, TJ, TM, TN, TR, TT, TZ, UA, UG, US, UZ, VC, VN, ZA, ZM, ZW. (30) Priority Data: 09159520.7 6 May 2009 (06.05.2009) EP (84) Designated States (unless otherwise indicated, for every kind of regional protection available): ARIPO (BW, GH, (71) Applicant (for all designated States except US): GM, KE, LR, LS, MW, MZ, NA, SD, SL, SZ, TZ, UG, NOVOZYMES A/S [DK/DK]; Krogshoejvej 36, ZM, ZW), Eurasian (AM, AZ, BY, KG, KZ, MD, RU, TJ, DK-2880 Bagsvaerd (DK). -

Environmental Properties of Chemicals Volume 2
1 t ENVIRONMENTAL 1 PROTECTION Esa Nikunen . Riitta Leinonen Birgit Kemiläinen • Arto Kultamaa Environmental properties of chemicals Volume 2 1 O O O O O O O O OO O OOOOOO Ol OIOOO FINNISH ENVIRONMENT INSTITUTE • EDITA Esa Nikunen e Riitta Leinonen Birgit Kemiläinen • Arto Kultamaa Environmental properties of chemicals Volume 2 HELSINKI 1000 OlO 00000001 00000000000000000 Th/s is a second revfsed version of Environmental Properties of Chemica/s, published by VAPK-Pub/ishing and Ministry of Environment, Environmental Protection Department as Research Report 91, 1990. The pubiication is also available as a CD ROM version: EnviChem 2.0, a PC database runniny under Windows operating systems. ISBN 951-7-2967-2 (publisher) ISBN 952-7 1-0670-0 (co-publisher) ISSN 1238-8602 Layout: Pikseri Julkaisupalvelut Cover illustration: Jussi Hirvi Edita Ltd. Helsinki 2000 Environmental properties of chemicals Volume 2 _____ _____________________________________________________ Contents . VOLUME ONE 1 Contents of the report 2 Environmental properties of chemicals 3 Abbreviations and explanations 7 3.1 Ways of exposure 7 3.2 Exposed species 7 3.3 Fffects________________________________ 7 3.4 Length of exposure 7 3.5 Odour thresholds 8 3.6 Toxicity endpoints 9 3.7 Other abbreviations 9 4 Listofexposedspecies 10 4.1 Mammais 10 4.2 Plants 13 4.3 Birds 14 4.4 Insects 17 4.5 Fishes 1$ 4.6 Mollusca 22 4.7 Crustaceans 23 4.8 Algae 24 4.9 Others 25 5 References 27 Index 1 List of chemicals in alphabetical order - 169 Index II List of chemicals in CAS-number order -

INDEX to PESTICIDE TYPES and FAMILIES and PART 180 TOLERANCE INFORMATION of PESTICIDE CHEMICALS in FOOD and FEED COMMODITIES
US Environmental Protection Agency Office of Pesticide Programs INDEX to PESTICIDE TYPES and FAMILIES and PART 180 TOLERANCE INFORMATION of PESTICIDE CHEMICALS in FOOD and FEED COMMODITIES Note: Pesticide tolerance information is updated in the Code of Federal Regulations on a weekly basis. EPA plans to update these indexes biannually. These indexes are current as of the date indicated in the pdf file. For the latest information on pesticide tolerances, please check the electronic Code of Federal Regulations (eCFR) at http://www.access.gpo.gov/nara/cfr/waisidx_07/40cfrv23_07.html 1 40 CFR Type Family Common name CAS Number PC code 180.163 Acaricide bridged diphenyl Dicofol (1,1-Bis(chlorophenyl)-2,2,2-trichloroethanol) 115-32-2 10501 180.198 Acaricide phosphonate Trichlorfon 52-68-6 57901 180.259 Acaricide sulfite ester Propargite 2312-35-8 97601 180.446 Acaricide tetrazine Clofentezine 74115-24-5 125501 180.448 Acaricide thiazolidine Hexythiazox 78587-05-0 128849 180.517 Acaricide phenylpyrazole Fipronil 120068-37-3 129121 180.566 Acaricide pyrazole Fenpyroximate 134098-61-6 129131 180.572 Acaricide carbazate Bifenazate 149877-41-8 586 180.593 Acaricide unclassified Etoxazole 153233-91-1 107091 180.599 Acaricide unclassified Acequinocyl 57960-19-7 6329 180.341 Acaricide, fungicide dinitrophenol Dinocap (2, 4-Dinitro-6-octylphenyl crotonate and 2,6-dinitro-4- 39300-45-3 36001 octylphenyl crotonate} 180.111 Acaricide, insecticide organophosphorus Malathion 121-75-5 57701 180.182 Acaricide, insecticide cyclodiene Endosulfan 115-29-7 79401 -

Method 8141B
METHOD 8141B ORGANOPHOSPHORUS COMPOUNDS BY GAS CHROMATOGRAPHY 1.0 SCOPE AND APPLICATION 1.1 This method provides procedures for the gas chromatographic (GC) determination of organophosphorus (OP) compounds. The compounds listed in the table below can be determined by GC using capillary columns with a flame photometric detector (FPD) or a nitrogen-phosphorus detector (NPD). Triazine herbicides can also be determined with this method when the NPD is used. Although performance data are presented for each of the listed chemicals, it is unlikely that all of them could be determined in a single analysis. This limitation results because the chemical and chromatographic behavior of many of these chemicals can result in co-elution. The analyst must select columns, detectors, and calibration procedures for the specific analytes of interest. Any listed chemical is a potential method interference when it is not a target analyte. Analyte CAS Registry No. Organophosphorus Pesticides Asponb 3244-90-4 Azinphos-methyl 86-50-0 Azinphos-ethyla 2642-71-9 Bolstar (Sulprofos) 35400-43-2 Carbophenothiona 786-19-6 Chlorfenvinphosa 470-90-6 Chlorpyrifos 2921-88-2 Chlorpyrifos methyla 5598-13-0 Coumaphos 56-72-4 Crotoxyphosa 7700-17-6 Demeton-Oc 8065-48-3 Demeton-Sc 8065-48-3 Diazinon 333-41-5 Dichlorofenthiona 97-17-6 Dichlorvos (DDVP) 62-73-7 Dicrotophosa 141-66-2 Dimethoate 60-51-5 Dioxathiona,c 78-34-2 Disulfoton 298-04-4 EPN 2104-64-5 Ethiona 563-12-2 Ethoprop 13194-48-4 Famphura 52-85-7 8141B - 1 Revision 2 November 2000 Analyte CAS Registry No. Fenitrothiona -

Organophosphate Poisoning : a Review
120 Sinha and Sharma Med J Indones Organophosphate poisoning : A review Parmod K. Sinha, Ashok Sharma Abstrak Pestisida organofosfat digunakan secara luas di seluruh dunia. Keracunan oleh bahan ini merupakan masalah kesehatan masyarakat, terutama di negara berkembang. Zat neurotoksik organofosfat merupakan bahan yang dianggap mengancam dalam bidang militer dan terorisme. Mekanisme toksisitas bahan ini adalah dengan cara menghambat asetilkolinesterase yang mengakibatkan menumpuknya neurotransmitor asetilkolin dan terjadi rangsangan terus-menerus pada reseptor asetilkolin pada sistem saraf sentral maupun perifer. Selain krisis kolinergik, organofosfat dapat menimbulkan berbagai sindrom neurologis, baik akut maupun kronik. Sedangkan gejala peralihan ( intermediate) terjadi 1-4 hari setelah krisis kolinergik teratasi. Pengobatan standar terdiri dari reaktivasi asetilkolinesterase dengan antidot golongan oksim (prolidoksim, oksidoksime, HI-6 dan HLo7), dan pengendalian efek biokimia asetilkolin dengan menggunakan atropin. Golongan oksim yang baru HI-6 dan Hlo7 merupakan reaktivator asetilkolinesterase yang lebih cocok dan efektif untuk keracunan akut dan berat dibandingkan dengan prolidoksim dan obidoksim. Penderita yang mendapat pengobatan segera, biasanya dapat sembuh dari toksisitas akut, namun gejala neurologis ikutan dapat saja terjadi. (Med J Indones 2003; 12: 120-6) Abstract Organophosphate pesticides are used extensively worldwide, and poisoning by these agents, particularly in developing nations is a public health problem. Organophosphorous -

Index to the NLM Classification 2011
National Library of Medicine Classification 2011 Index Disease see Tyrosinemias 1-8 5,12-diHETE see Leukotriene B4 1,2-Benzopyrones see Coumarins 5,12-HETE see Leukotriene B4 1,2-Dibromoethane see Ethylene Dibromide 5-HT see Serotonin 1,8-Dihydroxy-9-anthrone see Anthralin 5-HT Antagonists see Serotonin Antagonists 1-Oxacephalosporin see Moxalactam 5-Hydroxytryptamine see Serotonin 1-Propanol 5-Hydroxytryptamine Antagonists see Serotonin Organic chemistry QD 305.A4 Antagonists Pharmacology QV 82 6-Mercaptopurine QV 269 1-Sar-8-Ala Angiotensin II see Saralasin 7S RNA see RNA, Small Nuclear 1-Sarcosine-8-Alanine Angiotensin II see Saralasin 8-Hydroxyquinoline see Oxyquinoline 13-cis-Retinoic Acid see Isotretinoin 8-Methoxypsoralen see Methoxsalen 15th Century History see History, 15th Century 8-Quinolinol see Oxyquinoline 16th Century History see History, 16th Century 17 beta-Estradiol see Estradiol 17-Ketosteroids WK 755 A 17-Oxosteroids see 17-Ketosteroids A Fibers see Nerve Fibers, Myelinated 17th Century History see History, 17th Century Aardvarks see Xenarthra 18th Century History see History, 18th Century Abate see Temefos 19th Century History see History, 19th Century Abattoirs WA 707 2',3'-Cyclic-Nucleotide Phosphodiesterases QU 136 Abbreviations 2,4-D see 2,4-Dichlorophenoxyacetic Acid Chemistry QD 7 2,4-Dichlorophenoxyacetic Acid General P 365-365.5 Organic chemistry QD 341.A2 Library symbols (U.S.) Z 881 2',5'-Oligoadenylate Polymerase see Medical W 13 2',5'-Oligoadenylate Synthetase By specialties (Form number 13 in any NLM -

Review of Azinphos-Methyl Was Undertaken by the Office of Chemical Safety (OCS), Which Considered All the Toxicological Data and Information Submitted for the Review
The reconsideration of the active constituent azinphos-methyl, registrations of products containing azinphos-methyl and approvals of their associated labels PRELIMINARY REVIEW FINDINGS Volume 1: Review Summary OCTOBER 2006 Canberra Australia Azinphos-methyl review – Preliminary Review Findings © Australian Pesticides & Veterinary Medicines Authority 2006 This work is copyright. Apart from any use permitted under the Copyright Act 1968, no part may be reproduced without permission from the Australian Pesticides & Veterinary Medicines Authority. The Australian Pesticides & Veterinary Medicines Authority publishes this preliminary review findings report for the active constituent azinphos-methyl and products containing azinphos- methyl. For further information about this review or the Pesticides Review Program, contact: Manager Chemical Review Australian Pesticides & Veterinary Medicines Authority PO Box E 240 KINGSTON ACT 2604 Australia Telephone: 61 2 6272 3213 Facsimile: 61 2 6272 3218 Email: [email protected] APVMA web site: http://www.apvma.gov.au i Azinphos-methyl review – Preliminary Review Findings FOREWORD The Australian Pesticides & Veterinary Medicines Authority (APVMA) is an independent statutory authority with responsibility for the regulation of agricultural and veterinary chemicals in Australia. Its statutory powers are provided in the Agvet Codes scheduled to the Agricultural and Veterinary Chemicals Code Act 1994. The APVMA can reconsider the approval of an active constituent, the registration of a chemical product or -

2018 Treated Water Undetected Chemical Contaminant List
2018 Treated Water Undetected Chemical Contaminant List ESTROGENS AND OTHER HORMONES Diethylstilbestrol (DES) Estrone 17alpha-Estradiol 17alpha-Ethynal estradiol 17beta-Estradiol Progesterone Estriol cis-Testosterone trans-Testosterone INORGANIC CHEMICALS Antimony Niobium Arsenic Osmium Beryllium Palladium Cadmium Platinum Cerium Praseodymium Cesium Rhenium Cobalt Rhodium Cyanide Ruthenium Dysprosium Samarium Erbium Selenium Europium Silver Gadolinium Tantalum Gallium Tellurium Germanium Thallium Gold Thorium Hafnium Thulium Holmium Tin Iridium Titanium Lanthanum Tungsten Lead Uranium Lutetium Vanadium Mercury Ytterbium Molybdenum Zinc Neodymium Zirconium Nickel NITROSAMINES N-Nitropyrrolidine (NPYR) N-Nitrosomorpholine (NMOR) N-Nitrosodi-N-butylamine (NDBA) N-Nitrosodiphenylamine (NDPhA) N-Nitrosodiethylamine (NDEA) N-Nitrosodi-N-propylamine (NDPA) N-Nitrosodimethylamine (NDMA) N-Nitrosomethylethylamine (NMEA) N-Nitrosopiperidine (NPIP) 1 ORGANIC CHEMICALS Acenaphthene Butylbenzylphthalate Acenaphthylene Butyraldehyde (Butanal) Acetaldehyde Carbaryl Acetochlor Carbofuran Acetone Carbon disulfide Acrylamide Carbophenothion Acrylonitrile Carbon tetrachloride Alachlor Carboxin Aldicarb (Temik) Chlordane Aldicarb sulfone Chlordane, alpha Aldicarb sulfoxide Chlordane, gamma Aldrin Chlorfenvinphos Allyl chloride Chloroacetonitrile Tert-Amyl Methyl ether Chlorobenzene Ametryn Chlorobenzilate Anilizine 2-Chlorobiphenyl Anthracene 1-Chlorobutane Aspon Chloroethane Atraton Chloromethane Atrazine Chloroneb Azinphos-ethyl Chloroprene Azinphos-methyl -
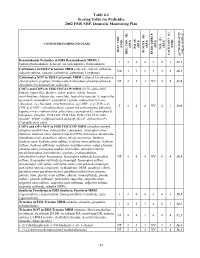
2002 NRP Section 6, Tables 6.1 Through
Table 6.1 Scoring Table for Pesticides 2002 FSIS NRP, Domestic Monitoring Plan } +1 0.05] COMPOUND/COMPOUND CLASS * ) (EPA) (EPA) (EPA) (EPA) (EPA) (FSIS) (FSIS) PSI (P) TOX.(T) L-1 HIST. VIOL. BIOCON. (B) {[( (2*R+P+B)/4]*T} REG. CON. (R) * ENDO. DISRUP. LACK INFO. (L) LACK INFO. {[ Benzimidazole Pesticides in FSIS Benzimidazole MRM (5- 131434312.1 hydroxythiabendazole, benomyl (as carbendazim), thiabendazole) Carbamates in FSIS Carbamate MRM (aldicarb, aldicarb sulfoxide, NA44234416.1 aldicarb sulfone, carbaryl, carbofuran, carbofuran 3-hydroxy) Carbamates NOT in FSIS Carbamate MRM (carbaryl 5,6-dihydroxy, chlorpropham, propham, thiobencarb, 4-chlorobenzylmethylsulfone,4- NT 4 1 3 NV 4 4 13.8 chlorobenzylmethylsulfone sulfoxide) CHC's and COP's in FSIS CHC/COP MRM (HCB, alpha-BHC, lindane, heptachlor, dieldrin, aldrin, endrin, ronnel, linuron, oxychlordane, chlorpyrifos, nonachlor, heptachlor epoxide A, heptachlor epoxide B, endosulfan I, endosulfan I sulfate, endosulfan II, trans- chlordane, cis-chlordane, chlorfenvinphos, p,p'-DDE, p, p'-TDE, o,p'- 3444NV4116.0 DDT, p,p'-DDT, carbophenothion, captan, tetrachlorvinphos [stirofos], kepone, mirex, methoxychlor, phosalone, coumaphos-O, coumaphos-S, toxaphene, famphur, PCB 1242, PCB 1248, PCB 1254, PCB 1260, dicofol*, PBBs*, polybrominated diphenyl ethers*, deltamethrin*) (*identification only) COP's and OP's NOT in FSIS CHC/COP MRM (azinphos-methyl, azinphos-methyl oxon, chlorpyrifos, coumaphos, coumaphos oxon, diazinon, diazinon oxon, diazinon met G-27550, dichlorvos, dimethoate, dimethoate -

Chemical Name Federal P Code CAS Registry Number Acutely
Acutely / Extremely Hazardous Waste List Federal P CAS Registry Acutely / Extremely Chemical Name Code Number Hazardous 4,7-Methano-1H-indene, 1,4,5,6,7,8,8-heptachloro-3a,4,7,7a-tetrahydro- P059 76-44-8 Acutely Hazardous 6,9-Methano-2,4,3-benzodioxathiepin, 6,7,8,9,10,10- hexachloro-1,5,5a,6,9,9a-hexahydro-, 3-oxide P050 115-29-7 Acutely Hazardous Methanimidamide, N,N-dimethyl-N'-[2-methyl-4-[[(methylamino)carbonyl]oxy]phenyl]- P197 17702-57-7 Acutely Hazardous 1-(o-Chlorophenyl)thiourea P026 5344-82-1 Acutely Hazardous 1-(o-Chlorophenyl)thiourea 5344-82-1 Extremely Hazardous 1,1,1-Trichloro-2, -bis(p-methoxyphenyl)ethane Extremely Hazardous 1,1a,2,2,3,3a,4,5,5,5a,5b,6-Dodecachlorooctahydro-1,3,4-metheno-1H-cyclobuta (cd) pentalene, Dechlorane Extremely Hazardous 1,1a,3,3a,4,5,5,5a,5b,6-Decachloro--octahydro-1,2,4-metheno-2H-cyclobuta (cd) pentalen-2- one, chlorecone Extremely Hazardous 1,1-Dimethylhydrazine 57-14-7 Extremely Hazardous 1,2,3,4,10,10-Hexachloro-6,7-epoxy-1,4,4,4a,5,6,7,8,8a-octahydro-1,4-endo-endo-5,8- dimethanonaph-thalene Extremely Hazardous 1,2,3-Propanetriol, trinitrate P081 55-63-0 Acutely Hazardous 1,2,3-Propanetriol, trinitrate 55-63-0 Extremely Hazardous 1,2,4,5,6,7,8,8-Octachloro-4,7-methano-3a,4,7,7a-tetra- hydro- indane Extremely Hazardous 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]- 51-43-4 Extremely Hazardous 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]-, P042 51-43-4 Acutely Hazardous 1,2-Dibromo-3-chloropropane 96-12-8 Extremely Hazardous 1,2-Propylenimine P067 75-55-8 Acutely Hazardous 1,2-Propylenimine 75-55-8 Extremely Hazardous 1,3,4,5,6,7,8,8-Octachloro-1,3,3a,4,7,7a-hexahydro-4,7-methanoisobenzofuran Extremely Hazardous 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)-carbonyl]oxime 26419-73-8 Extremely Hazardous 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)-carbonyl]oxime.