Trans-Andean Passage of Migrating Arctic Terns Over Patagonia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
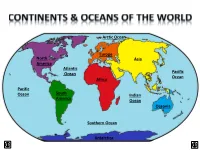
North America Other Continents
Arctic Ocean Europe North Asia America Atlantic Ocean Pacific Ocean Africa Pacific Ocean South Indian America Ocean Oceania Southern Ocean Antarctica LAND & WATER • The surface of the Earth is covered by approximately 71% water and 29% land. • It contains 7 continents and 5 oceans. Land Water EARTH’S HEMISPHERES • The planet Earth can be divided into four different sections or hemispheres. The Equator is an imaginary horizontal line (latitude) that divides the earth into the Northern and Southern hemispheres, while the Prime Meridian is the imaginary vertical line (longitude) that divides the earth into the Eastern and Western hemispheres. • North America, Earth’s 3rd largest continent, includes 23 countries. It contains Bermuda, Canada, Mexico, the United States of America, all Caribbean and Central America countries, as well as Greenland, which is the world’s largest island. North West East LOCATION South • The continent of North America is located in both the Northern and Western hemispheres. It is surrounded by the Arctic Ocean in the north, by the Atlantic Ocean in the east, and by the Pacific Ocean in the west. • It measures 24,256,000 sq. km and takes up a little more than 16% of the land on Earth. North America 16% Other Continents 84% • North America has an approximate population of almost 529 million people, which is about 8% of the World’s total population. 92% 8% North America Other Continents • The Atlantic Ocean is the second largest of Earth’s Oceans. It covers about 15% of the Earth’s total surface area and approximately 21% of its water surface area. -

South America Wine Cruise!
South America Wine Cruise! 17-Day Voyage Aboard Oceania Marina Santiago to Buenos Aires January 28 to February 14, 2022 Prepare to be awestruck by the magnificent wonders of South America! Sail through the stunning fjords of Patagonia and experience the cheerfully painted colonial buildings and cosmopolitan lifestyle of Uruguay and Argentina. Many people know about the fantastic Malbec, Torrontes, Tannat, and Carminiere wines that come from this area, but what they may not know is how many other great styles of wine are made by passionate winemakers throughout Latin America. This cruise will give you the chance to taste really remarkable wines from vineyards cooled by ocean breezes to those perched high in the snow-capped Andes. All made even more fun and educational by your wine host Paul Wagner! Your Exclusive Onboard Wine Experience Welcome Aboard Reception Four Exclusive Wine Paired Dinners Four Regional Wine Seminars Farewell Reception Paul Wagner Plus Enjoy: Renowned Wine Expert and Author Pre-paid Gratuities! (Expedia exclusive benefit!) "After many trips to Latin America, I want to share the wines, food and Complimentary Wine and Beer with lunch and dinner* culture of this wonderful part of the Finest cuisine at sea from Executive Chef Jacques Pépin world with you. The wines of these FREE Unlimited Internet (one per stateroom) countries are among the best in the Country club-casual ambiance world, and I look forward to Complimentary non-alcoholic beverages throughout the ship showing you how great they can be on this cruise.” *Ask how this can be upgraded to the All Inclusive Drink package onboard. -

New Siberian Islands Archipelago)
Detrital zircon ages and provenance of the Upper Paleozoic successions of Kotel’ny Island (New Siberian Islands archipelago) Victoria B. Ershova1,*, Andrei V. Prokopiev2, Andrei K. Khudoley1, Nikolay N. Sobolev3, and Eugeny O. Petrov3 1INSTITUTE OF EARTH SCIENCE, ST. PETERSBURG STATE UNIVERSITY, UNIVERSITETSKAYA NAB. 7/9, ST. PETERSBURG 199034, RUSSIA 2DIAMOND AND PRECIOUS METAL GEOLOGY INSTITUTE, SIBERIAN BRANCH, RUSSIAN ACADEMY OF SCIENCES, LENIN PROSPECT 39, YAKUTSK 677980, RUSSIA 3RUSSIAN GEOLOGICAL RESEARCH INSTITUTE (VSEGEI), SREDNIY PROSPECT 74, ST. PETERSBURG 199106, RUSSIA ABSTRACT Plate-tectonic models for the Paleozoic evolution of the Arctic are numerous and diverse. Our detrital zircon provenance study of Upper Paleozoic sandstones from Kotel’ny Island (New Siberian Island archipelago) provides new data on the provenance of clastic sediments and crustal affinity of the New Siberian Islands. Upper Devonian–Lower Carboniferous deposits yield detrital zircon populations that are consistent with the age of magmatic and metamorphic rocks within the Grenvillian-Sveconorwegian, Timanian, and Caledonian orogenic belts, but not with the Siberian craton. The Kolmogorov-Smirnov test reveals a strong similarity between detrital zircon populations within Devonian–Permian clastics of the New Siberian Islands, Wrangel Island (and possibly Chukotka), and the Severnaya Zemlya Archipelago. These results suggest that the New Siberian Islands, along with Wrangel Island and the Severnaya Zemlya Archipelago, were located along the northern margin of Laurentia-Baltica in the Late Devonian–Mississippian and possibly made up a single tectonic block. Detrital zircon populations from the Permian clastics record a dramatic shift to a Uralian provenance. The data and results presented here provide vital information to aid Paleozoic tectonic reconstructions of the Arctic region prior to opening of the Mesozoic oceanic basins. -

The Eu and the Arctic
MAGAZINE Dealing the Seal 8 No. 1 Piloting Arctic Passages 14 2016 THE CIRCLE The EU & Indigenous Peoples 20 THE EU AND THE ARCTIC PUBLISHED BY THE WWF GLOBAL ARCTIC PROGRAMME TheCircle0116.indd 1 25.02.2016 10.53 THE CIRCLE 1.2016 THE EU AND THE ARCTIC Contents EDITORIAL Leaving a legacy 3 IN BRIEF 4 ALYSON BAILES What does the EU want, what can it offer? 6 DIANA WALLIS Dealing the seal 8 ROBIN TEVERSON ‘High time’ EU gets observer status: UK 10 ADAM STEPIEN A call for a two-tier EU policy 12 MARIA DELIGIANNI Piloting the Arctic Passages 14 TIMO KOIVUROVA Finland: wearing two hats 16 Greenland – walking the middle path 18 FERNANDO GARCES DE LOS FAYOS The European Parliament & EU Arctic policy 19 CHRISTINA HENRIKSEN The EU and Arctic Indigenous peoples 20 NICOLE BIEBOW A driving force: The EU & polar research 22 THE PICTURE 24 The Circle is published quar- Publisher: Editor in Chief: Clive Tesar, COVER: terly by the WWF Global Arctic WWF Global Arctic Programme [email protected] (Top:) Local on sea ice in Uumman- Programme. Reproduction and 8th floor, 275 Slater St., Ottawa, naq, Greenland. quotation with appropriate credit ON, Canada K1P 5H9. Managing Editor: Becky Rynor, Photo: Lawrence Hislop, www.grida.no are encouraged. Articles by non- Tel: +1 613-232-8706 [email protected] (Bottom:) European Parliament, affiliated sources do not neces- Fax: +1 613-232-4181 Strasbourg, France. sarily reflect the views or policies Design and production: Photo: Diliff, Wikimedia Commonss of WWF. Send change of address Internet: www.panda.org/arctic Film & Form/Ketill Berger, and subscription queries to the [email protected] ABOVE: Sarek glacier, Sarek National address on the right. -

Endangered Andean Cat Distribution Beyond the Andes in Patagonia
original contribution ANDRES NOVARO1,2, SUSAN WALKER2*, ROCIO PALACIOS1,3, SEBASTIAN DI MARTINO4, MARTIN MONTEVERDE5, SEBASTIAN CANADELL6, LORENA RIVAS1,2 AND DANIEL COSSIOS7 Endangered Andean cat distribution beyond the Andes in Patagonia The endangered Andean cat Leopardus jacobita was considered an endemic of the Andes at altitudes above 3,000 m, until it was discovered in the Andean foothills of central Argentina in 2004. We carried out surveys for Andean cats and sympatric small cats in the central Andean foothills and the adjacent Patagonian steppe, and found Andean cats outside the Andes at elevations as low as 650 m. We determined that Andean cats are widespread but rare in the northern Patagonian steppe, with a patchy distribution. Our findings suggest that the species’ distribution may follow that of its principal prey, the rock-dwelling mountain vizcacha. The Andean cat was previously believed to be distribution if it does indeed follow that of the endemic to the Andes above 3,000 m (Yensen mountain vizcacha. First, to avoid bias for par- & Seymour 2000), until an opportunistic pho- ticular habitats beyond the Andes we placed Fig. 1. Location of new records and un- tograph in 2004 produced the startling finding a 2 x 2 km grid over the area with ArcGIS. confirmed reports of Andean cats in Men- of two Andean cats at only 1,800 m, in the We selected 105 grid cells, using stratified doza and Neuquén provinces (black dots), Andean foothills of central Argentina (Sorli et random sampling to ensure broad geographic relative to previous known distribution in al. -

Parasites of the Neotropic Cormorant Nannopterum (Phalacrocorax) Brasilianus (Aves, Phalacrocoracidae) in Chile
Original Article ISSN 1984-2961 (Electronic) www.cbpv.org.br/rbpv Parasites of the Neotropic cormorant Nannopterum (Phalacrocorax) brasilianus (Aves, Phalacrocoracidae) in Chile Parasitos da biguá Nannopterum (Phalacrocorax) brasilianus (Aves, Phalacrocoracidae) do Chile Daniel González-Acuña1* ; Sebastián Llanos-Soto1,2; Pablo Oyarzún-Ruiz1 ; John Mike Kinsella3; Carlos Barrientos4; Richard Thomas1; Armando Cicchino5; Lucila Moreno6 1 Laboratorio de Parásitos y Enfermedades de Fauna Silvestre, Departamento de Ciencia Animal, Facultad de Medicina Veterinaria, Universidad de Concepción, Chillán, Chile 2 Laboratorio de Vida Silvestre, Departamento de Ciencia Animal, Facultad de Medicina Veterinaria, Universidad de Concepción, Chillán, Chile 3 Helm West Lab, Missoula, MT, USA 4 Escuela de Medicina Veterinaria, Universidad Santo Tomás, Concepción, Chile 5 Universidad Nacional de Mar del Plata, Mar del Plata, Argentina 6 Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile How to cite: González-Acuña D, Llanos-Soto S, Oyarzún-Ruiz P, Kinsella JM, Barrientos C, Thomas R, et al. Parasites of the Neotropic cormorant Nannopterum (Phalacrocorax) brasilianus (Aves, Phalacrocoracidae) in Chile. Braz J Vet Parasitol 2020; 29(3): e003920. https://doi.org/10.1590/S1984-29612020049 Abstract The Neotropic cormorant Nannopterum (Phalacrocorax) brasilianus (Suliformes: Phalacrocoracidae) is widely distributed in Central and South America. In Chile, information about parasites for this species is limited to helminths and nematodes, and little is known about other parasite groups. This study documents the parasitic fauna present in 80 Neotropic cormorants’ carcasses collected from 2001 to 2008 in Antofagasta, Biobío, and Ñuble regions. Birds were externally inspected for ectoparasites and necropsies were performed to examine digestive and respiratory organs in search of endoparasites. -

Fastest Migration Highest
GO!” Everyone knows birds can fly. ET’S But not everyone knows that “L certain birds are really, really good at it. Meet a few of these champions of the skies. Flying Acby Ellen eLambeth; art sby Dave Clegg! Highest You don’t have to be a lightweight to fly high. Just look at a Ruppell’s griffon vulture (left). One was recorded flying at an altitude of 36,000 feet. That’s as high as passenger planes fly! In fact, it’s so high that you would pass out from lack of oxygen if you weren’t inside a plane. How does the vulture manage? It has Fastest (on the level) Swifts are birds that have that name for good special blood cells that make a small amount reason: They’re speedy! The swiftest bird using its own of oxygen go a long way. flapping-wing power is the common swift of Europe, Asia, and Africa (below). It’s been clocked at nearly 70 miles per hour. That’s the speed limit for cars on some highways. Vroom-vroom! Fastest (in a dive) Fastest Migration With gravity helping out, a bird can pick up extra speed. Imagine taking a trip of about 4,200 And no bird can go faster than a peregrine falcon in a dive miles. Sure, you could easily do it in an airplane. after prey (right). In fact, no other animal on Earth can go as But a great snipe (right) did it on the wing in just fast as a peregrine: more than 200 miles per hour! three and a half days! That means it averaged about 60 miles The prey, by the way, is usually another bird, per hour during its migration between northern which the peregrine strikes in mid-air with its balled-up Europe and central Africa. -

Climate, Tectonics, and the Morphology of the Andes
Climate, tectonics, and the morphology of the Andes David R. Montgomery Greg Balco Sean D. Willett Department of Geological Sciences, University of Washington, Seattle 98195-1310, USA ABSTRACT Large-scale topographic analyses show that hemisphere-scale climate variations are a ®rst-order control on the morphology of the Andes. Zonal atmospheric circulation in the Southern Hemisphere creates strong latitudinal precipitation gradients that, when incor- porated in a generalized index of erosion intensity, predict strong gradients in erosion rates both along and across the Andes. Cross-range asymmetry, width, hypsometry, and maximum elevation re¯ect gradients in both the erosion index and the relative dominance of ¯uvial, glacial, and tectonic processes, and show that major morphologic features cor- relate with climatic regimes. Latitudinal gradients in inferred crustal thickening and struc- tural shortening correspond to variations in predicted erosion potential, indicating that, like tectonics, nonuniform erosion due to large-scale climate patterns is a ®rst-order con- trol on the topographic evolution of the Andes. Keywords: geomorphology, erosion, tectonics, climate, Andes. INTRODUCTION we argue for the ®rst-order importance of earthquake cycle. Some studies have attribut- The presence or absence of mountain rang- large-scale climate zonations and resulting dif- ed local variations in structural, metamorphic, es at the global scale is determined by the lo- ferences in geomorphic processes to the mor- and geomorphic characteristics of the central cation and type of plate boundaries. Other fac- phology of mountain ranges. Andes to erosion (Gephart, 1994; Masek et al., tors become important in the evolution of 1994; Horton, 1999), but none has considered individual mountain systems. -

Roseate Tern Sterna Dougallii
COSEWIC Assessment and Update Status Report on the Roseate Tern Sterna dougallii in Canada Roseate Tern. Diane Pierce © 1995 ENDANGERED 2009 COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows: COSEWIC. 2009. COSEWIC assessment and update status report on the Roseate Tern Sterna dougallii in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vii + 48 pp. (www.sararegistry.gc.ca/status/status_e.cfm). Previous reports: COSEWIC. 1999. COSEWIC assessment and update status report on the Roseate Tern Sterna dougallii in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 28 pp. (www.sararegistry.gc.ca/status/status_e.cfm) Whittam, R.M. 1999. Update COSEWIC status report on the Roseate Tern Sterna dougallii in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-28 pp. Kirkham, I.R. and D.N. Nettleship. 1986. COSEWIC status report on the Roseate Tern Sterna dougallii in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 49 pp. Production note: COSEWIC would like to acknowledge Becky Whittam for writing the status report on the Roseate Tern Sterna dougallii in Canada, prepared under contract with Environment Canada, overseen and edited by Richard Cannings and Jon McCracken, Co-chairs, COSEWIC Birds Specialist Subcommittee. For additional copies contact: COSEWIC Secretariat c/o Canadian Wildlife Service Environment Canada Ottawa, ON K1A 0H3 Tel.: 819-953-3215 Fax: 819-994-3684 E-mail: COSEWIC/[email protected] http://www.cosewic.gc.ca Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur la Sterne de Dougall (Sterna dougallii) au Canada – Mise à jour. -

The Magic of Patagonia Holiday Grade
THE MAGIC OF Tour Code: 46300 Grade 5 PATAGONIA Holiday Duration: 15 nights HOLIDAY DATES Travelling through Chile and Argentina to 6th Nov 2019 - 21st Nov 2019 the end of the world... Walking in the awe-inspiring land of Patagonia, an area of uninterrupted lakes, dormant volcanoes and vast tracts of wilderness. HOLIDAY HIGHLIGHTS Admire the spectacular view and immense glaciers of Torres del Paine. See incredible wildlife - Guanacos, condors, rheas. Cross over the Andean watershed. Tango in Buenos Aires. THE MAGIC OF PATAGONIA HOLIDAY GRADE Where the craggy peaks of the Patagonian Andes This is a Grade 5 holiday. It combines walking in the are too steep sided to hold snow, bare rocks pierce Patagonian Andes with a little sightseeing in Buenos the sky. The famous Torres del Paine are perhaps Aires and Santiago, two iconic capital cities. There the most well known example of this. Snowy are plenty of opportunities for walking through mountains are common but here things are beautiful scenery. The walking in all of our different. Patagonian centres is not particularly high altitude. The highest point we reach on the holiday will likely be 1200m at Pliego Tumbado walking from El We begin our Patagonian journey in Buenos Aires Chalten. Paths are generally good underfoot. where we take in all the iconic sights and sounds of However they are sometimes stony and steep for the city, including an unmissable tango night. Then short sections so walking poles can be useful. on to El Chaten for some rewarding walking with magnificent views of Fitzroy and the surrounding peaks. -

A New Breeding Colony of Blue-Footed Booby Sula Nebouxii at Islote González, Archipelago Islas Desventuradas, Chile
Marín & González: A new Blue-footed Booby breeding colony in Chile 189 A NEW BREEDING COLONY OF BLUE-FOOTED BOOBY SULA NEBOUXII AT ISLOTE GONZÁLEZ, ARCHIPELAGO ISLAS DESVENTURADAS, CHILE MANUEL MARÍN1,2 & RODRIGO GONZÁLEZ3 1Natural History Museum of Los Angeles County, Section of Ornithology, 900 Exposition Boulevard, Los Angeles, California 90007, USA 2Current address: Casilla 15, Melipilla, Chile ([email protected]) 3Santa María 7178, Vitacura, Santiago, Chile Received 13 January 2021, accepted 02 February 2021 ABSTRACT MARÍN, M. & GONZÁLEZ, R. 2021. A new breeding colony of Blue-footed Booby Sula nebouxii at Islote González, Archipelago Islas Desventuradas, Chile. Marine Ornithology 49: 189–192. We describe a new disjunct population of Blue-footed Booby Sula nebouxii at Islote González in the Islas Desventuradas archipelago. Though likely much larger, the population size at the time of our survey was a minimum of 16 breeding pairs. On 14–15 December 2020, two pairs of boobies with nestlings and four pairs with empty nests were found. Likely the species is a recent arrival to the archipelago, perhaps during one of the strong El Niño events between 1970 and 2001 (especially 1982/83, when the species was first recorded in Chile). Blue-footed Booby might also nest on different areas at San Félix Island and some smaller islets around the archipelago, where they would have no nesting-site competition from the larger Masked Booby S. dactylatra. Key words: Blue-footed Booby, Sula nebouxii, breeding, Islas Desventuradas, Chile INTRODUCTION northern coast between 21°54′S and 23°05′S (Guerra 1983). Guerra observed 10 individuals in a 132-km transect while surveying for Within the Sulidae, a widespread and prolific group of piscivorous guano birds and fur seals along the coast. -

Why the Andes Matter
Sustainable Mountain Development RIO 2012 and beyond Why the Andes matter How the Andes contribute to sustainable development The Andes, covering a contiguous mountain region within Argentina, Bolivia, Chile, Colombia, Ecuador, Peru and Venezuela, occupy more than 2,500,000 km² and have The Andes, covering 33% of the area of the Ande- a population of about 85 million (45% of total country an countries, are vital for the livelihoods of the ma- populations), with the northern Andes as one of the most jority of the region’s population and the countries’ densely populated mountain regions in the world. At economies. However, increasing pressure, fuelled least a further 20 million people are also dependent on mountain resources and ecosystem services in the large by growing population numbers, changes in land cities along the Pacific coast of South America. use, unsustainable exploitation of resources, and climate change, could have far-reaching negati- The Andes play a vital part in national economies, ve impacts on ecosystem goods and services. To accounting for a significant proportion of the region’s achieve sustainable development, policy action is GDP, providing large agricultural areas, mineral resources, required regarding the protection of water resour- and water for agriculture, hydroelectricity (Figure 1), ces, responsible mining practices, adaptation to cli- domestic use, and some of the largest business centres in South America. However, some of the region’s poorest mate change and mechanisms to generate and use areas are also located in the mountains. knowledge for sound decision making. The region is highly diverse in terms of landscape, biodi- versity including agro-biodiversity, languages, peoples and cultures.