A NEW RECORD of the RHEOPHYTIC FERN Osmunda
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Growth of Fern Gametophytes After 20 Years of Storage in Liquid Nitrogen
FERN GAZ. 20(8): 337-346. 2018 337 GROWTH OF FERN GAMETOPHYTES AFTER 20 YEARS OF STORAGE IN LIQUID NITROGEN V. C. Pence Center for Conservation and Research of Endangered Wildlife (CREW) Cincinnati Zoo & Botanical Garden, 3400 Vine Street, Cincinnati, OH 45220, USA email: [email protected] Key words: cryopreservation, ex situ conservation, gametophyte; in vitro; long-term storage ABSTRACT In vitro grown gametophytes of six species of ferns, which had been cryopreserved using the encapsulation dehydration procedure, were evaluated for survival after 20 yrs of storage in liquid nitrogen. Tissues were rewarmed and transferred to a recovery medium with the same methods originally used to test pre-storage viability. All six species resumed growth. Post-storage viability was not consistently higher or lower than pre-storage viability of LN exposed tissues, likely reflecting the small sample sizes. However, these results demonstrate that long-term storage in liquid nitrogen is a viable option for preserving gametophytes of at least some fern species and could be utilized as an additional tool for preserving valuable gametophyte collections and for the ex situ conservation of fern biodiversity. INTRODUCTION For many species of ferns, gametophyte tissues have proven to be highly adaptable to growth in vitro (Table 1) . Most of these have been initiated through the aseptic germination of spores, although the aseptic germination of gemmae has also been demonstrated (Raine & Sheffield, 1997). As in vitro cultures, gametophytes can provide tissues for research and for propagation, both for ornamental ferns as well as for ferns of conservation concern. The ex situ conservation of ferns has traditionally relied on living collections and spore banks (Ballesteros, 2011). -

Millerocaulis Tekelili Sp. Nov., a New Species of Osmundalean Fern from the Aptian Cerro Negro Formation (Antarctica) Ezequiel I
This article was downloaded by: [INASP - Pakistan ] On: 28 July 2011, At: 10:58 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Alcheringa: An Australasian Journal of Palaeontology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/talc20 Millerocaulis tekelili sp. nov., a new species of osmundalean fern from the Aptian Cerro Negro Formation (Antarctica) Ezequiel I. Vera Available online: 27 Jul 2011 To cite this article: Ezequiel I. Vera (2011): Millerocaulis tekelili sp. nov., a new species of osmundalean fern from the Aptian Cerro Negro Formation (Antarctica), Alcheringa: An Australasian Journal of Palaeontology, DOI:10.1080/03115518.2011.576541 To link to this article: http://dx.doi.org/10.1080/03115518.2011.576541 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan, sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material. -

Palaeogeograph Y, Palaeoclimatology, Palaeoecology , 17(1975): 157--172 © Elsevier Scientific Publishing Company, Amsterdam -- Printed in the Netherlands
Palaeogeograph y, Palaeoclimatology, Palaeoecology , 17(1975): 157--172 © Elsevier Scientific Publishing Company, Amsterdam -- Printed in The Netherlands CLIMATIC CHANGES IN EASTERN ASIA AS INDICATED BY FOSSIL FLORAS. II. LATE CRETACEOUS AND DANIAN V. A. KRASSILOV Institute of Biology and Pedology, Far-Eastern Scientific Centre, U.S.S.R. Academy of Sciences, Vladivostok (U.S.S.R.) (Received June 17, 1974; accepted for publication November 11, 1974) ABSTRACT Krassilov, V. A., 1975. Climatic changes in Eastern Asia as indicated by fossil floras. II. Late Cretaceous and Danian. Palaeogeogr., Palaeoclimatol., Palaeoecol., 17:157--172. Four Late Cretaceous phytoclimatic zones -- subtropical, warm--temperate, temperate and boreal -- are recognized in the Northern Hemisphere. Warm--temperate vegetation terminates at North Sakhalin and Vancouver Island. Floras of various phytoclimatic zones display parallel evolution in response to climatic changes, i.e., a temperature rise up to the Campanian interrupted by minor Coniacian cooling, and subsequent deterioration of cli- mate culminating in the Late Danian. Cooling episodes were accompanied by expansions of dicotyledons with platanoid leaves, whereas the entire-margined leaf proportion increased during climatic optima. The floristic succession was also influenced by tectonic events, such as orogenic and volcanic activity which commenced in Late Cenomanian--Turonian times. Major replacements of ecological dominants occurred at the Maastrichtian/Danian and Early/Late Danian boundaries. INTRODUCTION The principal approaches to the climatic interpretation of fossil floras have been outlined in my preceding paper (Krassilov, 1973a). So far as Late Creta- ceous floras are concerned, extrapolation (i.e. inferences from tolerance ranges of allied modern taxa) is gaining in importance and the entire/non-entire leaf type ratio is no less expressive than it is in Tertiary floras. -

Hybrids in the Fern Genus Osmunda (Osmundaceae)
Bull. Natl. Mus. Nat. Sci., Ser. B, 35(2), pp. 63–69, June 22, 2009 Hybrids in the Fern Genus Osmunda (Osmundaceae) Masahiro Kato Department of Botany, National Museum of Nature and Science, Amakubo 4–1–1, Tsukuba, 305–0005 Japan E-mail address: [email protected] Abstract Four described putative hybrids in genus Osmunda, O. intermedia from Japan, O. rug- gii from eastern U.S.A., O. nipponica from central Japan, and O. mildei from southern China, are enumerated with notes on their hybridity. It is suggested that Osmunda intermedia is an intrasub- generic hybrid (O. japonica of subgenus Osmunda ϫ O. lancea of subgenus Osmunda), O. ruggii is an intersubgeneric hybrid (O. regalis of subgenus Osmunda ϫ O. claytoniana of subgenus Clay- tosmunda), O. nipponica is an intersubgeneric hybrid (O. japonica ϫ O. claytoniana of subgenus Claytosmunda), and O. midlei is an intersubgeneric hybrid (O. japonica ϫ O. angustifolia or O. vachellii of subgenus Plenasium). Among the four, O. intermedia is the most widely distributed and can reproduce in culture, suggesting that it can reproduce to some extent in nature. Key words : Hybrid, Osmunda intermedia, Osmunda mildei, Osmunda nipponica, Osmunda rug- gii. three subgenera Claytosmunda, Osmunda, and Introduction Plenasium, genus Leptopteris, genus Todea, and The genus Osmunda has been classified in ei- genus Osmundastrum (see also Metzgar et al., ther the broad or narrow sense. In the previously 2008). most accepted and the most lumping classifica- Four putative hybrids are known in the genus tion, it was divided into three subgenera, Osmun- Osmunda s.l. in eastern U.S.A. -

Plant Anatomy Lab 5
Plant Anatomy Lab 7 - Stems II This exercise continues the previous lab in studying primary growth in the stem. We will be looking at stems from a number of different plant species, and emphasize (1) the variety of stem tissue patterns, (2) stele types and the location of vascular tissues, (3) the development of the stem from meristem activity, and (4) the production of xylem and phloem by the procambium. All of the species studied are pictured either in your text or the atlases at the front of the lab. 1) Early vascular plants (cryptogams) A) Obtain a piece of the rachis of the fern (collected in the White Hall atrium). This structure is superficially analogous to a stem, although it has a different origin. Prepare a transverse section of the rachis and stain it in toluidine blue. Note the large amount of cortical tissue and the presence of sclerenchyma cells near the outer cortex. Also note that the stele vascular tissue appears to be amphiphloic. Lastly, you should be able to see readily the endodermal-style thickenings on the cells just outside each vascular bundle. B) Find prepared slides of the Osmunda and Polypodium (fern) rhizomes. Note the amphiphloic bundles (a protostele?), the presence of sclerenchyma in the cortex, and the thickened walls of the endodermis that you will find is not uncommon among many non-seed plants. Remember that rhizomes are modified stems that often occur underground. Osmunda fern vascular bundle. C) Obtain a prepared slide of Psilotum. This stele does not have a pith, so it is a protostele (or an actinostele because it has a star-like shape). -

Osmundaceae Royal Fern Family
Osmundaceae Royal Fern Family A family of large coarse plants, these are our largest pteridophyte species. Stipes arise from rhizomes Page | 70 covered in persistent frond bases. Stipes are pubescent but without scales, bearing stipules at their bases. Fertile fronds may be separate from vegetative fronds, or fronds divided between sterile and fertile portions. Sporangia are not arranged in sori, but in short stalked clusters. Osmunda L. royal ferns A single genus finds its way to eastern North America; three species grow in Nova Scotia. All have pinnate or bipinnate fronds. Key to species A. Fronds twice-pinnate; fertile pinnae carried at the top. Osmunda regalis aa. Fronds pinnate; sterile fronds deeply divided, lobes nearly entire. B B. Fertile pinnae near the middle of sterile fronds; no woolly O. claytoniana pubescence at the base of pinnae. bb. Fertile fronds on separate stipes; pinnae with a tuft of pubescence O. cinnamomea in the axils. Osmunda cinnamomea L. Cinnamon Fern; osmonde cannelle A common species, its fertile fronds are cinnamon– coloured, comprising minute sporangia appressed to the axis. Sterile fronds have pinnae subtended by clusters of pale pubescence. Fiddleheads emerge covered in pale or white coloured tomentum. Spores in spring. Grows in poorly-drained soils as found in bogs, swamps, coastal barrens and even low-lying pastures and wet conifer woods. 1-9 Osmundaceae Photo by Sean Blaney Very common throughout the province. Elsewhere from NF to ON, south to TX and FL; South America; Eurasia. Page | 71 Photo by David Mazerolle Osmunda claytoniana L. Interrupted Fern; osmonde de Clayton Fronds resemble the species above, but with both fertile and sterile pinnae on the same stipe in this species. -

The Ferns and Their Relatives (Lycophytes)
N M D R maidenhair fern Adiantum pedatum sensitive fern Onoclea sensibilis N D N N D D Christmas fern Polystichum acrostichoides bracken fern Pteridium aquilinum N D P P rattlesnake fern (top) Botrychium virginianum ebony spleenwort Asplenium platyneuron walking fern Asplenium rhizophyllum bronze grapefern (bottom) B. dissectum v. obliquum N N D D N N N R D D broad beech fern Phegopteris hexagonoptera royal fern Osmunda regalis N D N D common woodsia Woodsia obtusa scouring rush Equisetum hyemale adder’s tongue fern Ophioglossum vulgatum P P P P N D M R spinulose wood fern (left & inset) Dryopteris carthusiana marginal shield fern (right & inset) Dryopteris marginalis narrow-leaved glade fern Diplazium pycnocarpon M R N N D D purple cliff brake Pellaea atropurpurea shining fir moss Huperzia lucidula cinnamon fern Osmunda cinnamomea M R N M D R Appalachian filmy fern Trichomanes boschianum rock polypody Polypodium virginianum T N J D eastern marsh fern Thelypteris palustris silvery glade fern Deparia acrostichoides southern running pine Diphasiastrum digitatum T N J D T T black-footed quillwort Isoëtes melanopoda J Mexican mosquito fern Azolla mexicana J M R N N P P D D northern lady fern Athyrium felix-femina slender lip fern Cheilanthes feei net-veined chain fern Woodwardia areolata meadow spike moss Selaginella apoda water clover Marsilea quadrifolia Polypodiaceae Polypodium virginanum Dryopteris carthusiana he ferns and their relatives (lycophytes) living today give us a is tree shows a current concept of the Dryopteridaceae Dryopteris marginalis is poster made possible by: { Polystichum acrostichoides T evolutionary relationships among Onocleaceae Onoclea sensibilis glimpse of what the earth’s vegetation looked like hundreds of Blechnaceae Woodwardia areolata Illinois fern ( green ) and lycophyte Thelypteridaceae Phegopteris hexagonoptera millions of years ago when they were the dominant plants. -
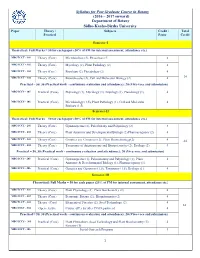
Syllabus for Post Graduate Course in Botany (2016 – 2017 Onward)
Syllabus for Post Graduate Course in Botany (2016 – 2017 onward) Department of Botany Sidho-Kanho-Birsha University Paper Theory / Subjects Credit / Total Practical Paper Credit Semester-I Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 101 Theory (Core) Microbiology (2), Phycology (2) 4 MBOTCCT - 102 Theory (Core) Mycology (2), Plant Pathology (2) 4 MBOTCCT - 103 Theory (Core) Bryology (2), Pteridology (2) 4 MBOTCCT - 104 Theory (Core) Biomolecules (2), Cell and Molecular Biology (2) 4 24 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 105 Practical (Core) Phycology (1), Mycology (1), Bryology (1), Pteridology (1). 4 MBOTCCS - 106 Practical (Core) Microbiology (1.5), Plant Pathology (1), Cell and Molecular 4 Biology (1.5). Semester-II Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 201 Theory (Core) Gymnosperms (2), Paleobotany and Palynology (2) 4 MBOTCCT - 202 Theory (Core) Plant Anatomy and Developmental Biology (2) Pharmacognosy (2) 4 MBOTCCT - 203 Theory (Core) Genetics and Genomics (2), Plant Biotechnology(2) 4 24 MBOTCCT - 204 Theory (Core) Taxonomy of Angiosperms and Biosystematics (2), Ecology (2) 4 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 205 Practical (Core) Gymnosperms (1), Palaeobotany and Palynology (1), Plant 4 Anatomy & Developmental Biology (1), Pharmacognosy (1). MBOTCCS - 206 Practical (Core) Genetics and Genomics (1.5), Taxonomy (1.5), Ecology (1). 4 Semester-III Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 301 Theory (Core) Plant Physiology (2), Plant Biochemistry (2) 4 MBOTCCT - 302 Theory (Core) Economic Botany (2), Bioinformatics (2) 4 MBOTCCT - 303 Theory (Core) Elements of Forestry (2), Seed Technology (2). -

Iop Newsletter 120
IOP NEWSLETTER 120 October 2019 CONTENTS Letter from the president Elections of IOP Executive Committee 2020 – Call for nominations Special issue in occasion of the 65th birthday of Hans Kerp Collection Spotlight: Cleveland Museum of Natural History Reflections from the ‘Earth Day’ 2019 Upcoming meetings IOP Logo: The evolution of plant architecture (© by A. R. Hemsley) 1 Letter from the president Greetings Members, This past quarter we have welcomed the publication Festschrifts celebrating our colleagues Hans Kerp (PalZ Paläontologische Zeitschrift, see below) and Gar Rothwell (International Journal of Plant Sciences v 180 nos. 7, 8). Thank you to the teams of editors and authors who have contributed to these issues illustrating the continuing strength of our discipline— and congratulations to Hans and Gar for inspiring such productivity by your admiring colleagues! Already three years have passed since our gathering for IOPC in Salvador, Brazil in 2016, and now planning is in full swing for next year’s meeting in Prague, 12-19 September 2020. This will be the 11th quadrennial meeting of IOP, held concurrently with the 15th International Palynological Conference. Please note the formal announcement in this newsletter (page 12, herein). Nominations for colleagues deserving of honorary membership are welcome at any time. As the submission of abstracts for IOPC/IPC presentations will be possible soon, we kindly remind the IOP Student Travel Awards. We will financially support about 5 to 7 PhD or MSc students in order to enable them to participate in the conference and present their research results in a talk. Recent PhD graduates will also qualify for these awards, if their completion was less than nine months prior to the time of the conference. -

Curriculum Vitae
CURRICULUM VITAE ORCID ID: 0000-0003-0186-6546 Gar W. Rothwell Edwin and Ruth Kennedy Distinguished Professor Emeritus Department of Environmental and Plant Biology Porter Hall 401E T: 740 593 1129 Ohio University F: 740 593 1130 Athens, OH 45701 E: [email protected] also Courtesy Professor Department of Botany and PlantPathology Oregon State University T: 541 737- 5252 Corvallis, OR 97331 E: [email protected] Education Ph.D.,1973 University of Alberta (Botany) M.S., 1969 University of Illinois, Chicago (Biology) B.A., 1966 Central Washington University (Biology) Academic Awards and Honors 2018 International Organisation of Palaeobotany lifetime Honorary Membership 2014 Fellow of the Paleontological Society 2009 Distinguished Fellow of the Botanical Society of America 2004 Ohio University Distinguished Professor 2002 Michael A. Cichan Award, Botanical Society of America 1999-2004 Ohio University Presidential Research Scholar in Biomedical and Life Sciences 1993 Edgar T. Wherry Award, Botanical Society of America 1991-1992 Outstanding Graduate Faculty Award, Ohio University 1982-1983 Chairman, Paleobotanical Section, Botanical Society of America 1972-1973 University of Alberta Dissertation Fellow 1971 Paleobotanical (Isabel Cookson) Award, Botanical Society of America Positions Held 2011-present Courtesy Professor of Botany and Plant Pathology, Oregon State University 2008-2009 Visiting Senior Researcher, University of Alberta 2004-present Edwin and Ruth Kennedy Distinguished Professor of Environmental and Plant Biology, Ohio -

Kasey Hartz Natural Area Reference Sheet Osmunda Cinnamomea Cinnamon Fern
Kasey Hartz Natural Area Reference Sheet Osmunda cinnamomea Cinnamon Fern Osmundaceae (Royal fern family) Blooming season: No flower but plant emerges in spring. Plant: A perennial plant that actively grows from spring to fall. Stipes are 10-20 cm long. They are pink to reddish brown with wooly hairs covering the stipe. The average mature height can reach 1.5 m. The rhizomes are ascending to creeping and form a large, vase like base. The minimum root depth is 30 cm. Leaves: Fronds appear as brownish-green fiddleheads covered with rusty wool in early spring. The fronds are dimorphic. The fertile fronds appear before the sterile fronds and look like a cinnamon stick, but are shorter than the sterile fronds. Oblong in shape; 15-45 cm long and 2-3 cm wide. 8-25 pairs of tripinnate, opposite oblong pinnae with groupings of brown sporangia. Sterile fronds are green, bipinnate, and form around the fertile ones. Elliptic in shape, 3-12 dm long, 13-30 cm wide, and acute at the tip. 15-25 opposite pinnae that have a tuft of red brown hair at the base of the pinna. Flower: No flower. Fruit: No fruit, but sporangia form spores on the fertile fronds. They are a brown cinnamon color. The spores mature May to June. Can be confused with: Royal fern because fronds have similar shapes, come from the same family, and both are typically found in the same area. Geographic range: Type specimen location: Creek Bed Habitat. State: Common throughout the whole of Michigan, but less common in the north. -

The Paraphyly of Osmunda Is Confirmed by Phylogenetic Analyses of Seven Plastid Loci
Systematic Botany (2008), 33(1): pp. 31–36 © Copyright 2008 by the American Society of Plant Taxonomists The Paraphyly of Osmunda is Confirmed by Phylogenetic Analyses of Seven Plastid Loci Jordan S. Metzgar,1 Judith E. Skog,2,3 Elizabeth A. Zimmer,4 and Kathleen M. Pryer1,5 1Department of Biology, Duke University, Durham, North Carolina 27708 U.S.A. 2Department of Environmental Science and Policy, George Mason University, Fairfax, Virginia 22030 U.S.A. 3National Science Foundation, Division of Biological Infrastructure, 4201 Wilson Blvd., Arlington, Virginia 22203 U.S.A. 4Department of Botany and Laboratory of Analytical Biology, Smithsonian Museum Support Center, 4210 Silver Hill Road, Suitland, Maryland 20746 U.S.A. 5Author for correspondence ([email protected]) Communicating Editor: Daniel Potter Abstract—To resolve phylogenetic relationships among all genera and subgenera in Osmundaceae, we analyzed over 8,500 characters of DNA sequence data from seven plastid loci (atpA, rbcL, rbcL–accD, rbcL–atpB, rps4–trnS, trnG–trnR, and trnL–trnF). Our results confirm those from earlier anatomical and single-gene (rbcL) studies that suggested Osmunda s.l. is paraphyletic. Osmunda cinnamomea is sister to the remainder of Osmundaceae (Leptopteris, Todea, and Osmunda s.s.). We support the recognition of a monotypic fourth genus, Osmundastrum, to reflect these results. We also resolve subgeneric relationships within Osmunda s.s. and find that subg. Claytosmunda is strongly supported as sister to the rest of Osmunda. A stable, well-supported classification for extant Osmundaceae is proposed, along with a key to all genera and subgenera. Keywords—ferns, Osmunda, Osmundaceae, Osmundastrum, paraphyly, plastid DNA.