And Ornithomimid (Theropoda)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Tooth Enamel Microstructure of Revueltosaurus
Palaeontologia Electronica palaeo-electronica.org Tooth enamel microstructure of Revueltosaurus and Krzyzanowskisaurus (Reptilia:Archosauria) from the Upper Triassic Chinle Group, USA: Implications for function, growth, and phylogeny Andrew B. Heckert and Jessica A. Miller-Camp ABSTRACT Tooth enamel microstructure can carry significant phylogenetic, ontogenetic, and functional information within amniotes. Here we provide the first descriptions of the tooth enamel microstructure of two Late Triassic taxa, the crurotarsan Revueltosaurus callenderi Hunt and the putative ornithischian Krzyzanowskisaurus hunti (Heckert), which some consider closely related. To test the hypotheses that enamel thickness corresponds to function and/or phylogeny we analyzed the enamel of each at various scales, measuring enamel thickness and examining microstructural features through- out both longitudinal and cross-sectional thickness using previously established tech- niques to facilitate comparisons. Both taxa possess thick (up to ~150 µm) enamel for their size (< 20 mm crown height). Enamel in R. callenderi ranged from ~5-152 µm across a premaxillary tooth in longitudinal section, and ~42-92 µm in a maxillary/den- tary tooth transverse section. K. hunti enamel thickness was ~18-155 µm longitudinally and ~29-75 µm transversely. Both also had well-developed basal unit layers (BUL) and weakly developed columnar microstructure. Well-developed lines of incremental growth (LIG) are present in both taxa, through which the columnar enamel grades into parallel crystallite enamel. Their enamel microstructure is therefore grossly similar to that of several ornithischian taxa, especially ankylosaurs, with which they are strongly convergent, and also compares well to rauisuchids and tyrannosaurids. The relatively unique combination of microstructural characteristics in the schmelzmuster of R. -

On the Presence of the Subnarial Foramen in Prestosuchus Chiniquensis (Pseudosuchia: Loricata) with Remarks on Its Phylogenetic Distribution
Anais da Academia Brasileira de Ciências (2016) (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201620150456 www.scielo.br/aabc On the presence of the subnarial foramen in Prestosuchus chiniquensis (Pseudosuchia: Loricata) with remarks on its phylogenetic distribution LÚCIO ROBERTO-DA-SILVA1,2, MARCO A.G. FRANÇA3, SÉRGIO F. CABREIRA3, RODRIGO T. MÜLLER1 and SÉRGIO DIAS-DA-SILVA4 ¹Programa de Pós-Graduação em Biodiversidade Animal, Universidade Federal de Santa Maria, Av. Roraima, 1000, Bairro Camobi, 97105-900 Santa Maria, RS, Brasil ²Laboratório de Paleontologia, Universidade Luterana do Brasil, Av. Farroupilha, 8001, Bairro São José, 92425-900 Canoas, RS, Brasil ³Laboratório de Paleontologia e Evolução de Petrolina, Campus de Ciências Agrárias, Universidade Federal do Vale do São Francisco, Rodovia BR 407, Km12, Lote 543, 56300-000 Petrolina, PE, Brasil 4Centro de Apoio à Pesquisa da Quarta Colônia, Universidade Federal de Santa Maria, Rua Maximiliano Vizzotto, 598, 97230-000 São João do Polêsine, RS, Brasil Manuscript received on July 1, 2015; accepted for publication on April 15, 2016 ABSTRACT Many authors have discussed the subnarial foramen in Archosauriformes. Here presence among Archosauriformes, shape, and position of this structure is reported and its phylogenetic importance is investigated. Based on distribution and the phylogenetic tree, it probably arose independently in Erythrosuchus, Herrerasaurus, and Paracrocodylomorpha. In Paracrocodylomorpha the subnarial foramen is oval-shaped, placed in the middle height of the main body of the maxilla, and does not reach the height of ascending process. In basal loricatans from South America (Prestosuchus chiniquensis and Saurosuchus galilei) the subnarial foramen is ‘drop-like’ shaped, the subnarial foramen is located above the middle height of the main body of the maxilla, reaching the height of ascending process, a condition also present in Herrerasaurus ischigualastensis. -

New Insights on Prestosuchus Chiniquensis Huene
New insights on Prestosuchus chiniquensis Huene, 1942 (Pseudosuchia, Loricata) based on new specimens from the “Tree Sanga” Outcrop, Chiniqua´ Region, Rio Grande do Sul, Brazil Marcel B. Lacerda1, Bianca M. Mastrantonio1, Daniel C. Fortier2 and Cesar L. Schultz1 1 Instituto de Geocieˆncias, Laborato´rio de Paleovertebrados, Universidade Federal do Rio Grande do Sul–UFRGS, Porto Alegre, Rio Grande do Sul, Brazil 2 CHNUFPI, Campus Amı´lcar Ferreira Sobral, Universidade Federal do Piauı´, Floriano, Piauı´, Brazil ABSTRACT The ‘rauisuchians’ are a group of Triassic pseudosuchian archosaurs that displayed a near global distribution. Their problematic taxonomic resolution comes from the fact that most taxa are represented only by a few and/or mostly incomplete specimens. In the last few decades, renewed interest in early archosaur evolution has helped to clarify some of these problems, but further studies on the taxonomic and paleobiological aspects are still needed. In the present work, we describe new material attributed to the ‘rauisuchian’ taxon Prestosuchus chiniquensis, of the Dinodontosaurus Assemblage Zone, Middle Triassic (Ladinian) of the Santa Maria Supersequence of southern Brazil, based on a comparative osteologic analysis. Additionally, we present well supported evidence that these represent juvenile forms, due to differences in osteological features (i.e., a subnarial fenestra) that when compared to previously described specimens can be attributed to ontogeny and indicate variation within a single taxon of a problematic but important -

Heptasuchus Clarki, from the ?Mid-Upper Triassic, Southeastern Big Horn Mountains, Central Wyoming (USA)
The osteology and phylogenetic position of the loricatan (Archosauria: Pseudosuchia) Heptasuchus clarki, from the ?Mid-Upper Triassic, southeastern Big Horn Mountains, Central Wyoming (USA) † Sterling J. Nesbitt1, John M. Zawiskie2,3, Robert M. Dawley4 1 Department of Geosciences, Virginia Tech, Blacksburg, VA, USA 2 Cranbrook Institute of Science, Bloomfield Hills, MI, USA 3 Department of Geology, Wayne State University, Detroit, MI, USA 4 Department of Biology, Ursinus College, Collegeville, PA, USA † Deceased author. ABSTRACT Loricatan pseudosuchians (known as “rauisuchians”) typically consist of poorly understood fragmentary remains known worldwide from the Middle Triassic to the end of the Triassic Period. Renewed interest and the discovery of more complete specimens recently revolutionized our understanding of the relationships of archosaurs, the origin of Crocodylomorpha, and the paleobiology of these animals. However, there are still few loricatans known from the Middle to early portion of the Late Triassic and the forms that occur during this time are largely known from southern Pangea or Europe. Heptasuchus clarki was the first formally recognized North American “rauisuchian” and was collected from a poorly sampled and disparately fossiliferous sequence of Triassic strata in North America. Exposed along the trend of the Casper Arch flanking the southeastern Big Horn Mountains, the type locality of Heptasuchus clarki occurs within a sequence of red beds above the Alcova Limestone and Crow Mountain formations within the Chugwater Group. The age of the type locality is poorly constrained to the Middle—early Late Triassic and is Submitted 17 June 2020 Accepted 14 September 2020 likely similar to or just older than that of the Popo Agie Formation assemblage from Published 27 October 2020 the western portion of Wyoming. -

Associated Skeletons of a New Middle Triassic “Rauisuchia” from Brazil
Naturwissenschaften (2011) 98:389–395 DOI 10.1007/s00114-011-0782-3 ORIGINAL PAPER Associated skeletons of a new middle Triassic “Rauisuchia” from Brazil Marco Aurélio G. França & Jorge Ferigolo & Max C. Langer Received: 28 January 2011 /Revised: 3 March 2011 /Accepted: 4 March 2011 /Published online: 29 March 2011 # Springer-Verlag 2011 Abstract For more than 30 million years, in early Mesozoic Introduction Pangea, “rauisuchian” archosaurs were the apex predators in most terrestrial ecosystems, but their biology and evolu- “Rauisuchians” are a widespread group of crocodile-line tionary history remain poorly understood. We describe a new archosaurs, first recognized by Huene (1942), based on “rauisuchian” based on ten individuals found in a single fossils collected early in the last century by Gilherme Rau locality from the Middle Triassic (Ladinian) Santa Maria in the Middle Triassic of Rio Grande do Sul, southern Formation of southern Brazil. Nine articulated and associat- Brazil. Since then, the group has been also recorded in Late ed skeletons were discovered, three of which have nearly Triassic and possibly Early Triassic rocks in all major parts complete skulls. Along with sedimentological and tapho- of Pangea, except Australia and Antarctica (Gower 2000; nomic data, this suggests that those highly successful Brusatte et al. 2010). With estimated lengths of up to 8 m predators exhibited some kind of intraspecific interaction. (Barberena 1978;Bonaparte1981), “rauisuchians” are Other monotaxic assemblages of Triassic archosaurs are Late generally regarded as the top terrestrial predators of their Triassic (Norian-Rhaetian) in age, approximately 10 million ecosystems. They are usually considered to be quadrupedal, years younger than the material described here. -

New Fossil Find in New Mexico Named After Artist Georgia O'keeffe 10 February 2006
New Fossil Find in New Mexico Named After Artist Georgia O'Keeffe 10 February 2006 modern crocodiles, Norell said. Obviously, this group of crocodiles and dinosaurs must have had similar habitats and probably fed in the same way, accounting for the similarities of the limbs and skull." Other examples of convergence include marsupial mammals related to kangaroos and opossums that evolved into creatures resembling lions and wolves. The features of Effigia okeeffeae also were Sketch courtesy of Sterling Nesbitt. unexpected. "It has large eyes, a beak, it walked on two feet and had small hands," Nesbitt said. Source: Columbia University Two Columbia scientists have discovered the fossil of a toothless crocodile relative that looks like a six- foot-long, two-legged dinosaur, but is actually a distant cousin of today's alligators and crocodiles. Adjunct professor of earth and environmental sciences Mark Norell and his graduate student Sterling Nesbitt, both of whom also work as paleontologists at the American Museum of Natural History, have named the fossil Effigia okeeffeae. Effigia means "ghost," referring to the decades that the fossil remained hidden from science. The species name, okeeffeae, honors the artist Georgia O'Keeffe, who lived near the site in northern New Mexico where the fossil was found. The discovery was announced last month in a technical paper in The Proceedings of the Royal Society and covered in the The New York Times. Click here to see. Scientists say Effigia is a striking example of "convergence," when two lineages evolve the same body plan. "It is astounding to see so many advanced dinosaur features in an animal so closely related to 1 / 2 APA citation: New Fossil Find in New Mexico Named After Artist Georgia O'Keeffe (2006, February 10) retrieved 1 October 2021 from https://phys.org/news/2006-02-fossil-mexico-artist-georgia-okeeffe.html This document is subject to copyright. -

Creature Descriptions
Creature Descriptions Coelophysis (Hollow form)- Height: 4 ft (1.2 m) Length: 9 ft (2.7 m) Weight: 100 lbs (45.5 kg) Coelophysis is one of the most primitive carnivorous dinosaurs and lived during the Late Triassic (230-200 mya). Hundreds of skeletons of Coelophysis have been discovered at Ghost Ranch, New Mexico -- one of the largest fossil deposits ever discovered. Coelophysis adults, young adults, and juveniles were all quickly buried together in a flash flood. The large range of ages gives scientists clues about how early dinosaurs grew. The large accumulation of Coelophysis also suggests that they lived and maybe even hunted in large packs. Coelophysis is a coelophysid, a group that is found all around the world. Coelophysis was lightly built and measured 6-10 feet in length. Many small, pointed teeth indicate that Coelophysis was a carnivore. During the Late Triassic, Coelophysis was not the dominant predator, but more likely the prey of rauisuchians such as Postosuchus or a phytosaur such as Smilosuchus. However, the shape and characteristics of the skeleton of Coelophysis will later be characteristic of some of the largest predators ever to walk the Earth. Confuciusornis (Confucius bird)- About the size of a crow, Length: 8 to 12 in (.2 to .3 m) Weight: 1 lbs (.45 kg) Confuciusornis is one of the oldest birds that lived during the Early Cretaceous (120 mya) in eastern Asia. It is one of the most common fossil birds from the Early Cretaceous -- hundreds of specimens ranging from juveniles to adults have been discovered. Confuciusornis indicates that birds rapidly diversified after their origin. -
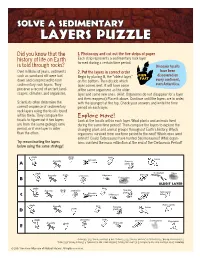
Solve a Sedimentary Layers Puzzle
SOLVE A SEDIMENTARY LAYERS PUZZLE Did you know that the 1. Photocopy and cut out the fi ve strips of paper history of life on Earth Each strip represents a sedimentary rock layer formed during a certain time period. is told through rocks? Dinosaur fossils Over millions of years, sediments 2. Put the layers in correct order have been such as sand and silt were laid FUN discovered on Begin by placing B, the “oldest layer,” FACT down and compressed to form on the bottom. Then decide which every continent, sedimentary rock layers. They layer comes next. It will have some even Antarctica. preserve a record of ancient land- of the same organisms as the older scapes, climates, and organisms. layer and some new ones. (Hint: Organisms do not disappear for a layer and then reappear.) Place it above. Continue until the layers are in order, Scientists often determine the with the youngest at the top. Check your answers and write the time correct sequence of sedimentary period on each layer. rock layers using the fossils found within them. They compare the Explore More! fossils to fi gure out if two layers Look at the fossils within each layer. What plants and animals lived are from the same geologic time during the same time period? Then compare the layers to explore the period, or if one layer is older changing plant and animal groups throughout Earth’s history. Which than the other. organisms survived from one time period to the next? Which ones went extinct? Could Tarbosaurus have hunted Seismosaurus? What organ- Try reconstructing the layers isms -

Triassic Vertebrate Paleontology in New Mexico
Archived version from NCDOCKS Institutional Repository http://libres.uncg.edu/ir/asu/ TRIASSIC VERTEBRATE PALEONTOLOGY IN NEW MEXICO By: Andrew B. Heckert and Spencer G. Lucas Abstract The Triassic vertebrate paleontological record of New Mexico includes important assemblages of tetrapod fossils from both the Middle Triassic Moenkopi Formation and the Upper Triassic Chinle Group. The Anton Chico Member of the Moenkopi Formation preserves primarily temnospondyl amphibian body fossils, but the record of reptiles comprises both sparse body fossil assemblages and more abundant track assemblages, mostly of chirotheriid reptiles. A bonebed accumulation of temnospondyls assigned to Eocyclotosaurus appetolatus is particularly notable. The Upper Triassic Chinle Group in New Mexico preserves an array of vertebrate faunal assemblages that represent the entirety of Chinle “time,” and includes numerous bonebeds of Revueltian age as well as the best records of Apachean vertebrates in the American West. These include the characteristic assemblages of the Revueltian and Apachean land- vertebrate faunachrons. Lucas, S. G. and Sullivan, R. M., eds. , 2015, Fossil Vertebrates in New Mexico. New Mexico Museum of Natural History and Science Bulletin 68. New Mexico Museum of Natural History and Science Lucas, S. G. and Sullivan, R. M., eds. , 2015, Fossil Vertebrates in New Mexico. New Mexico Museum of Natural History and Science Bulletin 68. 77 TRIASSIC VERTEBRATE PALEONTOLOGY IN NEW MEXICO ANDREW B. HECKERT1 and SPENCER G. LUCAS2 1Department of Geology, ASU Box 32067, Appalachian State University, Boone, NC 28608-2067 -email: [email protected]; 2New Mexico Museum of Natural History & Science, 1801 Mountain Road NW, Albuquerque, NM 87104 Abstract—The Triassic vertebrate paleontological record of New Mexico includes important assemblages of tetrapod fossils from both the Middle Triassic Moenkopi Formation and the Upper Triassic Chinle Group. -

An Enigmatic New Archosauriform from the Carnian– Norian, Upper Triassic, Ischigualasto Formation of Northwestern Argentina
Editors' choice An enigmatic new archosauriform from the Carnian– Norian, Upper Triassic, Ischigualasto Formation of northwestern Argentina IMANOL YÁÑEZ, DIEGO POL, JUAN MARTÍN LEARDI, OSCAR A. ALCOBER, and RICARDO N. MARTÍNEZ Yáñez, I., Pol, D., Leardi, J.M., Alcober, O.A., and Martínez, R.N. 2021. An enigmatic new archosauriform from the Carnian–Norian, Upper Triassic, Ischigualasto Formation of northwestern Argentina. Acta Palaeontologica Polonica 66 (X): xxx–xxx. In this contribution we introduce a new Late Triassic archosaur, Incertovenator longicollum gen. et sp. nov., with an unusual combination of character states that are present in certain early avemetatarsalian and pseudosuchian archosaur clades. The holotype consists of a partial postcranial skeleton, preserving most of the axial skeleton and displaying a marked anteroposterior elongation in the cervical vertebrae. We include I. longicollum gen. et sp. nov. into one of the most comprehensive early archosaur phylogenetic data sets available, and recover it as either an early diverging avemetatarsalian, closely associated with the clade Aphanosauria and Ornithodira, or as an early diverging loricatan closely related to Mandasuchus tanyauchen in the most parsimonious trees. We further evaluate which alternative phylogenetic positions can I. longicollum gen. et sp. nov. take in the suboptimal trees, and determined which character states support those alternative positions in comparison with those of the unconstrained analysis. The analyses recover the new taxon in three main general phylogenetic placements within Archosauria, as well as one position outside this clade, highlighting widespread morphological evolutionary convergence towards neck elongation in several clades of Triassic archosauriforms. Key words: Archosauria, Archosauriformes, Ischigualasto, osteology, phylogeny, taxonomy, Late Triassic, Argentina. -

The Late Triassic Ischigualasto Formation at Cerro Las Lajas
www.nature.com/scientificreports OPEN The Late Triassic Ischigualasto Formation at Cerro Las Lajas (La Rioja, Argentina): fossil tetrapods, high‑resolution chronostratigraphy, and faunal correlations Julia B. Desojo1,2*, Lucas E. Fiorelli1,3, Martín D. Ezcurra1,4, Agustín G. Martinelli1,4, Jahandar Ramezani5, Átila. A. S. Da Rosa6, M. Belén von Baczko1,4, M. Jimena Trotteyn1,7, Felipe C. Montefeltro8, Miguel Ezpeleta1,9 & Max C. Langer10 Present knowledge of Late Triassic tetrapod evolution, including the rise of dinosaurs, relies heavily on the fossil‑rich continental deposits of South America, their precise depositional histories and correlations. We report on an extended succession of the Ischigualasto Formation exposed in the Hoyada del Cerro Las Lajas (La Rioja, Argentina), where more than 100 tetrapod fossils were newly collected, augmented by historical fnds such as the ornithosuchid Venaticosuchus rusconii and the putative ornithischian Pisanosaurus mertii. Detailed lithostratigraphy combined with high‑precision U–Pb geochronology from three intercalated tufs are used to construct a robust Bayesian age model for the formation, constraining its deposition between 230.2 ± 1.9 Ma and 221.4 ± 1.2 Ma, and its fossil-bearing interval to 229.20 + 0.11/− 0.15–226.85 + 1.45/− 2.01 Ma. The latter is divided into a lower Hyperodapedon and an upper Teyumbaita biozones, based on the ranges of the eponymous rhynchosaurs, allowing biostratigraphic correlations to elsewhere in the Ischigualasto‑Villa Unión Basin, as well as to the Paraná Basin in Brazil. The temporally calibrated Ischigualasto biostratigraphy suggests the persistence of rhynchosaur-dominated faunas into the earliest Norian. Our ca. 229 Ma age assignment to Pi. -

Online Supplementary Appendices Appendix S1
ONLINE SUPPLEMENTARY APPENDICES APPENDIX S1: DESCRIPTION OF NEW CHARACTERS Note: characters numbered according to their location in the full analysis. 9. Supratemporal fenestra oriented primarily dorsally (0), mostly dorsally but visible as a sliver in lateral view (1), or extensively laterally (2). The supratemporal fenestrae of most archosaur outgroups (e.g. Erythrosuchus: Gower 2003; proterochampsids: Sill 1967, Romer 1971c, Arcucci 1990) face completely dorsally, a condition also seen in basal dinosaurs (Fig. 3B; Galton 1984, 1985b; Colbert 1989; Sereno & Novas 1993), phytosaurs (Chatterjee 1978), ornithosuchids (Walker 1964; Bonaparte 1971), and many “rauisuchians” (Arizonasaurus: Nesbitt 2005; Batrachotomus: Fig. 3F, Gower 1999; Effigia: Nesbitt 2007; Postosuchus: Long & Murry 1995; Shuvosaurus: Chatterjee 1993). In contrast, the supratemporal fenestrae of most aetosaurs face fully laterally (Fig. 3C), a condition often held as an autapomorphy of the clade (e.g. Heckert & Lucas 1999). This state is also seen in Gracilisuchus (MCZ 4116, 4117; Romer 1972a; Brinkman 1981). Additionally, it is apparent that many crurotarsan taxa possess supratemporal fenestrae that are partially exposed laterally, and thus visible as a narrow sliver in lateral view (Erpetosuchus: Benton & Walker 2002; Lotosaurus: IVPP unnumbered; Prestosuchus: Fig. 3E, Barberena 1978; Saurosuchus: Alcober 2000; Teratosaurus: Sulej 2005; Tikisuchus: Chatterjee & Majumdar 1987; basal crocodylomorphs: Terrestrisuchus, Sphenosuchus, Crush, 1984 Walker 1990). In many “rauisuchian” taxa this appears to be at least partially the result of dorsolateral rotation of the squamosal. Whereas in most archosaurs (e.g. Batrachotomus: SMNS 80260) the 1 ventral ramus of the squamosal projects straight ventrally, in “rauisuchians” with a slightly laterally facing fenestra (e.g. Rauisuchus: BPSG AS XXV 62) the ventral ramus projects lateroventrally, and thus the primitive “lateral” surface faces partially dorsally.