Steady-State Visually Evoked Potential Correlates of Object Recognition Memory
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bach in the Subways
TO SEE The best gigs in L.A. this season hometown show in support of its JANUARY latest dancy EP. à Lodge Room, 104 N Ave 56 (323-509- 2861, lodgeroomhlp.com). Feb 9 at 8pm; $15. Wolf Parade Fresh off an arena tour with Cuco Arcade Fire, the veteran indie- The local teenage heartthrob rock act headlines in support of takes his laid-back synths, its latest well-received record. Chicano roots and bilingual à The FondA TheAtre, 6126 Hollywood Blvd rhymes from small clubs to (323-464-6269, fondatheatre.com). Jan 19, this theater-sized stage. DON’T MISS 20 at 9pm; $35. à El Rey TheAtre, 5515 WIlshIre Blvd (323-936-6400, theelrey.com). Feb 14 Ty Segall at 9pm; $20. The tireless singer-songwriter Bach in the Subways takes the stage to celebrate The Piano Recital the release of his latest Listen to intimate solo piano YOU EXPECT DELAYS and elbow-to-elbow crowds on fuzzed-out retro-rock record, compositions played by the the subway. You don’t expect to hear Bach’s Cello Suites. Freedom’s Goblin. likes of the Hush Sound’s Greta “I was looking around thinking, Man, people never hear à Teragram Ballroom, 1234 W 7th St (213- Morgan, Guster’s Ryan Miller, this music,” says Dale Henderson. So in 2010, he decided 689-9100, terAgrAmbAllroom.com). Fruit Bats’ Eric D. Johnson to remedy that problem by the most efficient means he Jan 26–28 at 9pm; $26. and soulful singer-songwriter could imagine: by playing the cello on New York City Kevin Morby. -

Songs by Title
Karaoke Song Book Songs by Title Title Artist Title Artist #1 Nelly 18 And Life Skid Row #1 Crush Garbage 18 'til I Die Adams, Bryan #Dream Lennon, John 18 Yellow Roses Darin, Bobby (doo Wop) That Thing Parody 19 2000 Gorillaz (I Hate) Everything About You Three Days Grace 19 2000 Gorrilaz (I Would Do) Anything For Love Meatloaf 19 Somethin' Mark Wills (If You're Not In It For Love) I'm Outta Here Twain, Shania 19 Somethin' Wills, Mark (I'm Not Your) Steppin' Stone Monkees, The 19 SOMETHING WILLS,MARK (Now & Then) There's A Fool Such As I Presley, Elvis 192000 Gorillaz (Our Love) Don't Throw It All Away Andy Gibb 1969 Stegall, Keith (Sitting On The) Dock Of The Bay Redding, Otis 1979 Smashing Pumpkins (Theme From) The Monkees Monkees, The 1982 Randy Travis (you Drive Me) Crazy Britney Spears 1982 Travis, Randy (Your Love Has Lifted Me) Higher And Higher Coolidge, Rita 1985 BOWLING FOR SOUP 03 Bonnie & Clyde Jay Z & Beyonce 1985 Bowling For Soup 03 Bonnie & Clyde Jay Z & Beyonce Knowles 1985 BOWLING FOR SOUP '03 Bonnie & Clyde Jay Z & Beyonce Knowles 1985 Bowling For Soup 03 Bonnie And Clyde Jay Z & Beyonce 1999 Prince 1 2 3 Estefan, Gloria 1999 Prince & Revolution 1 Thing Amerie 1999 Wilkinsons, The 1, 2, 3, 4, Sumpin' New Coolio 19Th Nervous Breakdown Rolling Stones, The 1,2 STEP CIARA & M. ELLIOTT 2 Become 1 Jewel 10 Days Late Third Eye Blind 2 Become 1 Spice Girls 10 Min Sorry We've Stopped Taking Requests 2 Become 1 Spice Girls, The 10 Min The Karaoke Show Is Over 2 Become One SPICE GIRLS 10 Min Welcome To Karaoke Show 2 Faced Louise 10 Out Of 10 Louchie Lou 2 Find U Jewel 10 Rounds With Jose Cuervo Byrd, Tracy 2 For The Show Trooper 10 Seconds Down Sugar Ray 2 Legit 2 Quit Hammer, M.C. -

Songs by Artist
Songs by Artist Title Title (Hed) Planet Earth 2 Live Crew Bartender We Want Some Pussy Blackout 2 Pistols Other Side She Got It +44 You Know Me When Your Heart Stops Beating 20 Fingers 10 Years Short Dick Man Beautiful 21 Demands Through The Iris Give Me A Minute Wasteland 3 Doors Down 10,000 Maniacs Away From The Sun Because The Night Be Like That Candy Everybody Wants Behind Those Eyes More Than This Better Life, The These Are The Days Citizen Soldier Trouble Me Duck & Run 100 Proof Aged In Soul Every Time You Go Somebody's Been Sleeping Here By Me 10CC Here Without You I'm Not In Love It's Not My Time Things We Do For Love, The Kryptonite 112 Landing In London Come See Me Let Me Be Myself Cupid Let Me Go Dance With Me Live For Today Hot & Wet Loser It's Over Now Road I'm On, The Na Na Na So I Need You Peaches & Cream Train Right Here For You When I'm Gone U Already Know When You're Young 12 Gauge 3 Of Hearts Dunkie Butt Arizona Rain 12 Stones Love Is Enough Far Away 30 Seconds To Mars Way I Fell, The Closer To The Edge We Are One Kill, The 1910 Fruitgum Co. Kings And Queens 1, 2, 3 Red Light This Is War Simon Says Up In The Air (Explicit) 2 Chainz Yesterday Birthday Song (Explicit) 311 I'm Different (Explicit) All Mixed Up Spend It Amber 2 Live Crew Beyond The Grey Sky Doo Wah Diddy Creatures (For A While) Me So Horny Don't Tread On Me Song List Generator® Printed 5/12/2021 Page 1 of 334 Licensed to Chris Avis Songs by Artist Title Title 311 4Him First Straw Sacred Hideaway Hey You Where There Is Faith I'll Be Here Awhile Who You Are Love Song 5 Stairsteps, The You Wouldn't Believe O-O-H Child 38 Special 50 Cent Back Where You Belong 21 Questions Caught Up In You Baby By Me Hold On Loosely Best Friend If I'd Been The One Candy Shop Rockin' Into The Night Disco Inferno Second Chance Hustler's Ambition Teacher, Teacher If I Can't Wild-Eyed Southern Boys In Da Club 3LW Just A Lil' Bit I Do (Wanna Get Close To You) Outlaw No More (Baby I'ma Do Right) Outta Control Playas Gon' Play Outta Control (Remix Version) 3OH!3 P.I.M.P. -
CURATED and HOSTED by MATT LAROCCA “FORTY SIX & 2” Tool, Arr
in partnership with SATURDAY FEBRUARY 27, 2021 AT 7:30PM ARTSRIOT, BURLINGTON CURATED AND HOSTED BY MATT LAROCCA “FORTY SIX & 2” Tool, arr. Vitamin String Quartet An intricate and rhythmically explosive mixed-meter song off the second album released by Tool in 1996. “IF YOU NEED ME” Kat Wright, arr. Matt LaRocca “If You Need Me” is a song that will be released on Kat’s next album, reimagined with a string quartet style reminiscent of George Gershwin. “DIDO’S LAMENT” Henry Purcell, arr. Matt LaRocca Purcell’s 1689 opera Dido and Aeneas contains the impossibly MATT LAROCCA, beautiful and heartbreaking aria, “Dido’s Lament.” Kat lends her unique voice and sound to this operatic classic. CREATIVE PROJECTS CHAIR “PARANOID ANDROID” Matt LaRocca is a composer, performer, Radiohead, arr. Eric Gorfain and educator who you are just as likely “Paranoid Android” is the lead single from Radiohead’s 1997 to see playing with a band in a dive Grammy-winning album OK Computer. Radiohead is often bar as conducting an orchestra. He is regarded as the most influential musical group since the Beatles. on the composition and theory faculty “VOODOO DOLLS” at the University of Vermont, and is Jessie Montgomery the Artistic Director of the Champlain Jessie Montgomery is an acclaimed composer, violinist, and Philharmonic Orchestra. He is also the the recipient of the Leonard Bernstein Award from the ASCAP Executive Director of Music-COMP, an Foundation. “Voodoo Dolls” was commissioned by the JUMP! Dance Company of Rhode Island and is influenced by West organization that teaches composition African drumming patterns and chant motives. -

~Üchs\~T~Tische Berichte Des Statistischen Landesamtes
r . ' ' ~ "\..., 0taüsns~~:es Lcnaesan .~ Schiesvvig-l·ioistein !. ~üchs\~t~tische Berichte des Statistischen Landesamtes Schleswig- Holstein ~ Kiel, Fröbelstraße 15 -17 ~ Postanschrift Postfach 1141, 2300 Kiell ~ Fernruf: (0431) 6 89 51 Nachdruck, auch auszugsweise, ist gern gestattet, wenn die Quelle genannt wird Stati sti sehe Berichte mit * vor der Nummerung enthalten Angaben, die olle Stati sti sehen Landesämter für ihren Bereich unter gl eieher Kennzifferveröffentlichen G IV 1 - j/81~ 82 und 83 Ausgegeben am 7. 11. 1984 Der Fremdenverkehr in den Gemeinden Schleswig-Holsteins 1981, 1982 und 1983 Zuständiger Dezernent: König Der vorliegende Statistische Bericht enthält Zahlen über den Fremdenverkehr in Schleswig-Holstein für die Kalenderjahre 1981, 1982 und 1983 nach Gemeinden und Kreisen. Die Angaben beziehen sich auf Seherbergungsstätten mit 9 und mehr Betten (einschließlich Jugendherbergen) sowie auf Campingplätze mit 6 und mehr Stellplätzen. Neben den Eckdaten des Fremdenverkehrs, der Zahl der Betriebe, Betten (nur für Beherbergungsstätten), Gäste und Übernachtungen w~rden je Ge meinde die Verweildauer der Gäste und die Zahl der Übernachtungen je Einwohner nachgewiesen, ferner für die Seherbergungsstätten die Zahl der Übernachtungen je Bett, für die Campingplätze der Anteil der auf die Dauercamper entfallenden Übernachtungen sowie der Anteil der Übernachtungen auf Campingplätzen an den Übernachtungen der Seherbergungsstätten und Campingplätzen zusammen. Die Seher bergungsstätten und die Campingplätze sind nach der Systematik der Wirtschafts zweige, Ausgabe 1979 abgegrenzt worden. Zeltlager werden folglich bei den Be herbergungsstätten nachgewiesen. Ergebnisse für Gemeinden mit weniger als drei Betrieben bzw. Campingplätzen unter liegen der statistischen Geheimhaltung. In den Tabellen sind die betreffenden Felder gepunktet. Die Kreissummen und die Landessummen sind jedoch vollständig. -
No Ordinary World Duran Duran’S Simon Le Bon Hungry to Keep Audiences Dancing Zoe-Ruth Photography
VOLUME 10, NUMBER 5 NUMBER 10, VOLUME NO ORDINARY WORLD DURAN DURAN’S SIMON LE BON HUNGRY TO KEEP AUDIENCES DANCING ZOE-RUTH PHOTOGRAPHY 418 Sheridan Road Highland Park, IL 60035 847-266-5000 www.ravinia.org Welz Kauffman President and CEO Nick Pullia Communications Director, Executive Editor Nick Panfl Publications Manager, Editor Alexandra Pikeas Graphic Designer IN THIS ISSUE FEATURES DEPARTMENTS Since 1991 12 Practice Room with a View 9 Message from the Chairman 3453 Commercial Ave., Northbrook, IL 60062 Te Juilliard Quartet’s Joseph Lin and and President www.performancemedia.us | 847-770-4620 Astrid Schween prepare musicians for 31 Rewind Gail McGrath - Publisher & President more than the concert stage. Sheldon Levin - Publisher & Director of Finance By Wynne Delacoma 50 Ravinia’s Steans Music Institute 18 Grateful Eternal Account Managers 52 Reach*Teach*Play Jerry Garcia and the Grateful Dead Rand Brichta - Arnie Hoffman - Greg Pigott made music for all time. 57 Salute to Sponsors Southwest By Davis Schneiderman Betsy Gugick & Associates 972-387-1347 24 No Ordinary World 73 Annual Fund Donors Duran Duran’s Simon Le Bon is hungry Sales & Marketing Consultant 80 Corporate Partners to keep audiences dancing. Mike Hedge 847-770-4643 81 Corporate Matching Gifs David L. Strouse, Ltd. 847-835-5197 By Miriam Di Nunzio 34 ‘Home Schooled’ 82 Special Gifs Cathy Kiepura - Graphic Designer Jonathan Biss and Pamela Frank recall Lory Richards - Graphic Designer their master classes in growing up as 83 Event Sponsors A.J. Levin - Director of Operations (and with) musicians. 84 Board of Trustees Josie Negron - Accounting By Mark Tomas Ketterson 85 Women’s Board Joy Morawez - Accounting 40 Unplugged, Unbound Willie Smith - Supervisor Operations Chris Cornell fnds ‘higher truth’ 86 Associates Board Earl Love - Operations in his music by going acoustic. -
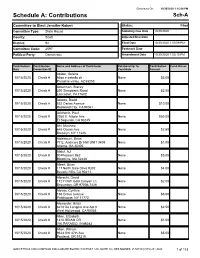
Schedule A: Contributions Sch-A
Generated On: 10/30/2020 1:15:04 PM Schedule A: Contributions Sch-A Committee to Elect Jennifer Kakert Status: Filed Committee Type: State House Statutory Due Date 10/30/2020 County: Scott Adjusted Due Date District: 92 Filed Date 10/30/2020 1:00:09 PM Committee Code: 2597 Postmark Date Political Party: Democratic Amendment Date 10/30/2020 1:00:15 PM Contribution Contribution Name and Address of Contributor Relationship To Contribution Fund-Raiser Date Committee ID Candidate Amount Abidor, Selena 10/15/2020 Check # 66oo e caballo dr None $5.00 Paradise valley, AZ 85253 Ackerman, Stacey 10/15/2020 Check # 290 Steepbank Road None $2.50 Lancaster, PA 17602 Adams, David 10/15/2020 Check # 522 Carlos Avenue None $10.00 Redwood City, CA 94061 Adelstein, Paul 10/15/2020 Check # 2260 E. Maple Ave. None $50.00 El Segundo, CA 90245 Ahl, Matthew 10/15/2020 Check # 645 Ocean Ave. None $2.50 Brooklyn, NY 11226 Aidelbaum, Brian 10/15/2020 Check # 77 E. Andrews Dr NW UNIT 3409 None $1.00 Atlanta, GA 30305 Alaluf, AJ 10/15/2020 Check # 80 Pleasant #62 None $5.00 Brookline, MA 02446 Albert, Brian 10/15/2020 Check # 117 North Gale Drive #302 None $4.00 Beverly Hills, CA 90211 Albrecht, David 10/15/2020 Check # 17211 NW Gold Canyon Ln None $2.00 Beaverton, OR 97006-7426 Alevas, Cynthia 10/15/2020 Check # 166 Grove avenue None $5.00 Patchogue, NY 11772 Alexander, Brian 10/15/2020 Check # 8218 De Longpre Ave Apt 9 None $2.50 West Hollywood, CA 90046 Allen, Elizabeth 10/15/2020 Check # 1810 BRIAM DR None $1.00 VALPARAISO, IN 46383 Allen, William 10/15/2020 -
Pops, Guster Make Quite an Edgy Team
MUSIC REVIEW Pops, Guster make quite an edgy team By Joan Anderman, Globe Staff | June 23, 2005 Beer and roadies under the gilded Beethoven insignia topped the list of firsts at Symphony Hall last night, where the Boston Pops kicked off its groundbreaking Pops on the Edge series with the alternative pop band Guster. Additional oddities included girls rushing the stage, whoops loud enough to peel the paint off the statues, and the sight of Keith Lockhart pogo-ing on the podium. If the series' goal is to lure a younger crowd to the hallowed hall -- mission accomplished. University of Connecticut students Sean Morton and Kristen Kelleher -- both 23 and fretting that they were underdressed -- said they were excited to be attending their first classical music concert. It was with them and the throngs of other flip-flopped, jeans-clad audience members in mind that Lockhart programmed the first half of the evening with ''the music of revolution." The Danse sacral from Stravinsky's 1913 ''The Rite of Spring" was so controversial in its day, Lockhart explained, that at the premiere the dancers couldn't hear the musicians over the furor in the hall. Ninety-two years later it's as noisy and jagged as a rock tune. Full appreciation of the harmonic structure in Debussy's ''Prelude to the Afternoon of a Faun" required a subtler grasp of musical revolutions, but the work's liquid beauty was within everyone's reach. ''Totally amazing" is how one 13-year-old girl assessed Tod Machover's computer-enhanced, video-augmented ''Jeux Deux" for hyperpiano and orchestra -- a wild, lovely, and utterly modern collision of art and science. -

Folge 35/1986
35. Folge Hohe Ehrung für Werner Lippke Be der Sitzung der Ostpreiißischen Landesvertretung am 3. und 4. Mai 1986 n RotenburgIWumme wurde unser Kreisaitester Werner Lippke vom Sprecher unserer Landsmannschaft. Staatssekretär Dr Ottfried Henng, lur sene Verdien- ste um unsere Heimat mit der goldenen Ehrennadel der Landsmannschaft aus^ gezeichnet. Werner Lippke ist Mitglied der Kreisgemeinschaft Wehlau und damit der Landsmannschaft seit deren GrUndung. Er war von 1968 bs 1978 Kresver- treter. Er legte seinerzeit dies Amt aus gesundhetlichen Grunden nieder. nimmt aber von 1978 an bis heute das Amt des Kreisaltesten (Vorsitzender des Kreis- tages) wahr Während seiner Zet als Kreisvertreter hat er U. a. es mUglich gemacht. da8 das lange geplante Hematbuch des Kreises Wehau endlich 1975 erschien. Vor zwei Jahren hat er - nachdem die Frage der Unterbringung des Hematmu- seums Wehlau in Syke gekart war - die vorhandenen Bestande geordnet. regi- striert und das Museum eingerichtet. so daß cs ]ctzt der Offentlichkeit Zugang- Iich ist. Er bearbeitet seit Jahren das Bildarchv unseres Kreses. das Archiv und die Bücherei. Während seiner Tatiaket als Rektor einer Sonderschule hat er den Kndern - so weit es in diesem Rahmen mogch war - Ostdeutschland und besonders Ostpreußen nahe gebracht. Im Werkunterricht entstanden viele Mo- dele ostpreußischer Burgen. Kirchen und Rathäuser. die bei Ausstellungen der Schule qezeigt wurden und so auch die Eltern ansprachen. Seit einigen Jahren ist ~ernerGppke Ortsvorsitzender des Bundes der vertriebenen-n senem Wohnort Kaltenkrchen Mit seinen Lichtbdvortragen uber Ostpreußen tragt er dazu bei daß bei Vertriebenen und Einheimischen das Bewußtsen fur die Be- deutung unserer Heimat lebendig bebt Werner Lippke hat sich um die Heimat verdient gemacht und tut es auch weiterhin noch rm Umschlag, vorne, innen Unser Titelbild: Schloß Ripkeim, erbaut 1791. -

The Fellows of the American Bar Foundation
THE FELLOWS OF THE AMERICAN BAR FOUNDATION 2011 Chair 2011-2012 Doreen D. Dodson Chair – Elect Myles V. Lynk Secretary Don Slesnick The Fellows is an honorary organization of attorneys, judges and law professors whose profes- sional, public and private careers have demonstrated outstanding dedication to the welfare of their communities and to the highest principles of the legal profession. Established in 1955, The Fellows encourage and support the research program of the American Bar Foundation. The American Bar Foundation works to advance justice through research on law, legal institutions, and legal processes. Current research covers such topics as end-of-life decision making, the value of early childhood education, how lawyers in public interest law organizations conceptualize and pursue their goals, what people think of the civil justice system against the backdrop of the politics of tort reform and the changes in the law that have resulted from the tort reform movement, and the factors that play a psychological role in laypersons’ decisions about justice and responsibility. The Fellows of the American Bar Foundation 750 N. Lake Shore Drive, 4th Floor Chicago, IL 60611 (800) 292-5065 Fax: (312) 988-6579 [email protected] www.americanbarfoundation.org OFFICERS AND DIRECTORS OF THE OFFICERS OF THE FELLOWS, CONT’D AMERICAN BAR FOUNDATION Don Slesnick, Secretary William C. Hubbard, President Slesnick & Casey LLP Hon. Bernice B. Donald, Vice President 2701 Ponce De Leon Boulevard, Suite 200 David A. Collins, Treasurer Coral Gables, FL 33134-6041 Ellen J. Flannery, Secretary Office: (305) 448-5672 Robert L. Nelson, ABF Director Fax: (305) 448-5685 Susan Frelich Appleton [email protected] Mortimer M. -

The Difference
The Difference In 1934, Muzak opened its doors and the business music industry was born. Today, satellite radio provides their music programs to businesses as well. But that’s where the similarities end. Remember how your mom used to tell you, “never judge a book by its cover”? She was right, you know. Satellite radio may have programs that look similar to ours, but dig deeper. Beyond the surface, the differences are striking. The Difference: Our Program “It’s more than just putting a few songs together... it’s Audio Architecture.” To show the amount of thought and passion that goes into our music programs, we asked Audio Architect Dean Carlson to talk about how he created Backpages, one of our core programs. How Dean Describes Backpages Backpages is a hip and dynamic music program featuring three basic music elements. At the core of the program, you will hear the biggest artists in the adult alternative world, such as Sheryl Crow, John Mayer and Tom Petty. We also feature alternative bands like U2, R.E.M. and Guster, who focus more on songwriting and unique instrumentation. Finally, you will hear some of the more up-and-coming artists of the genre like David Gray, Jack Johnson and Norah Jones. The result is a diverse and intelligent collection of both familiar and soon-to- be-discovered acoustic-based rock music. But what really sets Backpages apart is the attention given to each song’s selection and the flow between tracks. When you listen to our program compared to the competition, the difference is obvious. -

Volume 3 – Anthroponomastics
‘Names and Their Environment’ Proceedings of the 25th International Congress of Onomastic Sciences Glasgow, 25-29 August 2014 Volume 3 Anthroponomastics Edited by Carole Hough Daria Izdebska University of Glasgow Glasgow 2016 ISBN 10: 0-85261-947-2 (for a set of five volumes) ISBN 13: 978-0-85261-947-6 The articles in this publication are © 2016 with the individual authors. They are made freely available under the terms of the Creative Commons licence (CC BY-NC-ND 4.0). For details, see: http://creativecommons.org/licenses/by-nc-nd/4.0/ Table of Contents (Volume 3) Balodis, Pauls (Latvia) Latvian Surnames Motivated by Profession ............................................................................ 1 Bichlmeier, Harald (Deutschland) Zum Anteil der Familiennamen slawischer Herkunft im deutschen Familiennamenschatz ............................................................................................................. 11 Brylla, Eva (Sweden) A New Personal Names Act in Sweden? ............................................................................... 27 Corino Rovano, Silvia (Italy) One Knight, Several Names ................................................................................................... 34 Demsky, Aaron (Israel) New Hebrew Names in the Second Temple Period: A By-Product of Biblical Exegesis ..... 44 Dräger, Kathrin (Deutschland) Abbe bis Zyprian. Patronyme im Deutschen Familiennamenatlas (abstract) ........................ 52 Eskola, Ksenia (Finland) Name Giving of Russian-Finnish Families (abstract)