Phenotypic Diversity Assessment Wihin a Major Ex Situ Collection of Coffees
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Endemic Chromoblastomycosis Caused Predominantly by Fonsecaea Nubica, Madagascar
Article DOI: https://doi.org/10.3201/eid2606.191498 Endemic Chromoblastomycosis Caused Predominantly by Fonsecaea nubica, Madagascar Appendix Appendix Table. Characteristics of clinical isolates from patients with suspected chromoblastomycosis in Madagascar and reference strains used in this study* Panfugal PCR for S/C Specific PCR for S/C MALDI-ToF GenBank accession no. Ccar- Code Species for D1D2, ITS Source Origin NL-1/NL-4 ITS1/ITS4 F/Ccar-R Fon-F/Fon-R MSP ID MYC04002 Unidentified NA, NA Biopsy Vatovavy Fitovinany N/P P/N N/N N/NA NA NA MYC04004 Fonsecaea sp. NA, NA Biopsy Vatovavy Fitovinany P/P P/NA N/NA N/P NA NA MYC08006 Fonsecaea sp. MK828325, NA Biopsy Analanjirofo P/P P/NA N/N N/P NA NA MYC10010 Fonsecaea sp. NA, NA Biopsy Melaky P/P P/NA N/NA P/P NA NA MYC10011 Fonsecaea sp. NA, NA Squama Vatovavy Fitovinany P/P P/NA N/NA P/P NA NA MYC10014 Cladophialophora carrionii MK828362, MK820046 Squama Androy P/P P/P N/P N/N X NA MYC03022 Unidentified NA, NA Biopsy Sofia NA/NA NA/NA N/NA NA/NA NA NA MYC04026 Fonsecaea nubica MK828326, MK828125 Biopsy Alaotra Mangoro N/P N/P NA/N NA/P NA X MYC05032 F. nubica MK828327, MK828124 Squama Amoron I Mania N/P N/P NA/N NA/P NA NA MYC06033 Unidentified NA, NA Biopsy Androy N/P N/P NA/N NA/N NA NA MYC08040 C. carrionii MK828363, MK820047 Biopsy Atsimo Andrefana N/P N/P NA/P NA/N NA NA MYC09041 Unidentified NA, NA Biopsy Analamanga NA/NA NA/NA NA/NA NA NA NA MYC03048 Unidentified NA, NA Biopsy Vakinankaratra N/P N/N N/N N/N NA NA MYC06057 Unidentified NA, NA Biopsy Amoron I Mania P/P N/N N/N N/N NA NA MYC06059 F. -
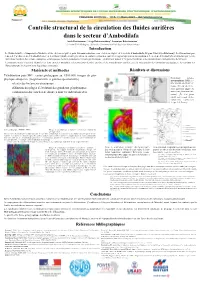
Contrôle Structural De La Circulation Des Fluides Aurifères Dans Le
Contrôle structural de la circulation des fluides aurifères dans le secteur d’Ambodilafa Anick Ratefiarimino 1, Serge Rakotonanahary1, Dominique Rakotomanana1 1 : Mention Génie Géologique (Université d’Antananarivo-Ecole Supérieure Polytechnique) Introduction Le District aurifère d’Ampasary—Vohilava est une des aires réputées pour leur minéralisation en or et au sein duquel est le secteur d’Ambodilafa (Région Vatovavy—Fitovinany). Les formations por- teuses de l’or du secteur d’Ambodilafa sont en des filons hydrothermaux généralement conformes dans des contextes magmatiques ou métamorphiques. Le secteur d’Ambodilafa est structuré par les dé- formations résultant des actions conjuguées et juxtaposées de deux événements majeurs gondwanéens : cisaillement majeur d’Angavo-Ifanadiana et les déformations curvi-planaires de Maroala. Les travaux menés visaient à identifier les liens entre les modalités et les processus de mise en place de la minéralisation aurifère avec la structuration des formations géologiques ; les conduits des fluides pouvant être façonnés par la géologie structurale. Matériels et méthodes Résultats et discussions Télédétection puis SIG : cartes géologiques au 1/500.000, images de géo- Evénement tectono- physique aéroportée (magnétométrie et gamma-spectrométrie) métamorphique DM1 : re- relevés des linéaments structuraux prise des plis antérieurs ver- ticaux (F1) par un événe- définition des plages d’évolution des grandeurs géophysiques ment postérieur auquel est associé un plissement sub- combinaison des couches de données dont les indications d’or vertical F2. Les plans axiaux sont sécants. Défor- mation finie : replissement de type 2 de Ramsay Carte géologique (PGRM, 2008) Image de géophysique aéroportée (ci-dessus): champ ma gnétique total. Lors de leur décantation les minéraux ont été orientés en fonction des caractères du champ magnétique terrestre régnant. -

RAPPORT D'activité 2015-2016 Projet D'adaptation De La Gestion Des Zones Côtières Au Changement Climatique
17' 0( (/ 1( ¶( 1 & 2 2 5 / , 2 9 * 1 , ( ( ¶ / ( 7 ( ' ' ( ( 6 5 ) ( 2 7 6 5 , ( 1 , 7 6 0 MINISTERE DE L’ENVIRONNEMENT, DE L’ECOLOGIE ET DES FORETS SECRETARIAT GENERAL BUREAU NATIONAL DE COORDINATION DES CHANGEMENTS CLIMATIQUES RAPPORT D'ACTIVITÉ 2015-2016 Projet d'Adaptation de la gestion des zones côtières au changement climatique PROJET D’AdaptatioN DE LA GESTION DES ZONES CÔTIÈRES AU CHANGEMENT CLIMatiQUE Etant un pays insulaire, Madagascar est Plusieurs actions ont été entreprises par le considéré comme l’un des pays les plus projet d’Adaptation de la gestion des Zones SOMMAIRE vulnérables à la variabilité et aux changements Côtières au changement climatique en tenant climatiques. Les dits changements se compte de l’Amélioration des écosystèmes CONTEXTE 5 manifestent surtout par le «chamboulement et des moyens de subsistance » au cours du régime des pluviométries, l’augmentation de l’année 2016 comme la réalisation des COMPOSANTE 1 : RENForcement DES capacITÉS de la température, la montée du niveau de études de vulnérabilité dans les quatre zones INSTITUTIONNELLES AUX Impacts DU CHANGEMENT la mer et l’intensification des évènements d’intervention, la création d’un mécanisme de CLImatIQUE DANS LES SITES DU proJET climatiques extrêmes tels que les cyclones, les coordination et la mise en place de la Gestion (MENABE, BOENY, VatovavY FItovINANY ET ATSINANANA) 7 inondations et les sècheresses. Devant cette Intégrée des zones côtières dans les régions situation alarmante, des actions d’adaptation Atsinanana, Boeny, et Vatovavy Fitovinany, ainsi COMPOSANTE 2 : RÉHABILItatION ET GESTION DES ZONES sont déja mises en oeuvre à Madagascar afin de que la mise en œuvre des scénarios climatiques CÔTIÈRES EN VUE d’uNE RÉSILIENCE À LONG TERME 17 renforcer la résilience de la population locale et à l’échelle réduite de ces quatre régions. -

Boissiera 71
Taxonomic treatment of Abrahamia Randrian. & Lowry, a new genus of Anacardiaceae BOISSIERA from Madagascar Armand RANDRIANASOLO, Porter P. LOWRY II & George E. SCHATZ 71 BOISSIERA vol.71 Director Pierre-André Loizeau Editor-in-chief Martin W. Callmander Guest editor of Patrick Perret this volume Graphic Design Matthieu Berthod Author instructions for www.ville-ge.ch/cjb/publications_boissiera.php manuscript submissions Boissiera 71 was published on 27 December 2017 © CONSERVATOIRE ET JARDIN BOTANIQUES DE LA VILLE DE GENÈVE BOISSIERA Systematic Botany Monographs vol.71 Boissiera is indexed in: BIOSIS ® ISSN 0373-2975 / ISBN 978-2-8277-0087-5 Taxonomic treatment of Abrahamia Randrian. & Lowry, a new genus of Anacardiaceae from Madagascar Armand Randrianasolo Porter P. Lowry II George E. Schatz Addresses of the authors AR William L. Brown Center, Missouri Botanical Garden, P.O. Box 299, St. Louis, MO, 63166-0299, U.S.A. [email protected] PPL Africa and Madagascar Program, Missouri Botanical Garden, P.O. Box 299, St. Louis, MO, 63166-0299, U.S.A. Institut de Systématique, Evolution, Biodiversité (ISYEB), UMR 7205, Centre national de la Recherche scientifique/Muséum national d’Histoire naturelle/École pratique des Hautes Etudes, Université Pierre et Marie Curie, Sorbonne Universités, C.P. 39, 57 rue Cuvier, 75231 Paris CEDEX 05, France. GES Africa and Madagascar Program, Missouri Botanical Garden, P.O. Box 299, St. Louis, MO, 63166-0299, U.S.A. Taxonomic treatment of Abrahamia (Anacardiaceae) 7 Abstract he Malagasy endemic genus Abrahamia Randrian. & Lowry (Anacardiaceae) is T described and a taxonomic revision is presented in which 34 species are recog- nized, including 19 that are described as new. -

Analysis of the Variability of Floral and Pollen Traits in Apple Cultivars—Selecting Suitable Pollen Donors for Cider Apple Orchards
agronomy Article Analysis of the Variability of Floral and Pollen Traits in Apple Cultivars—Selecting Suitable Pollen Donors for Cider Apple Orchards Alvaro Delgado 1,* , Muriel Quinet 2 and Enrique Dapena 1 1 Servicio Regional de Investigación y Desarrollo Agroalimentario (SERIDA), Apdo.13, E-33300 Villaviciosa, Asturias, Spain; [email protected] 2 Earth and Life Institute-Agronomy, Université Catholique de Louvain, Croix du Sud 4-5, Box L7 07 13, 1348 Louvain-la-Neuve, Belgium; [email protected] * Correspondence: [email protected] Abstract: Most apple trees (Malus domestica Borkh.) are self-incompatible and fruit yield depends on cross-pollination between genetically compatible cultivars with synchronous flowering. Flowering intensity can vary strongly among years due to the biennial bearing habit of the cultivars. The knowledge of the phenological stages and floral and pollen characteristics is essential to select suitable pollen donors. We evaluated the phenotypic variability of flowering-related traits (i.e., flowering phenology, flowering intensity, pollen production and pollen quality) in 45 apple cultivars over two successive flowering seasons. Large phenotypic variability was found among the studied cultivars indicating that the local germplasm collection provides a good source of genetic and phenotypic diversity. However, low correlations were observed between floral biology traits and, consequently, the improvement in one trait seems not to affect other traits. Some of the cultivars such as ‘Perurico’ and ‘Raxila Dulce’ regularly produced copious amounts of high-quality pollen Citation: Delgado, A.; Quinet, M.; which can improve the pollen load dispersion leading to a most effective pollination process. We did Dapena, E. Analysis of the Variability not identify statistically significant correlations between pollen attributes and the biennial bearing of Floral and Pollen Traits in Apple phenomenon. -

Pollinators in Forests – an Annotated Bibliography
Pollinators in Forests – An Annotated Bibliography This annotated bibliography was compiled while researching pollinators in the woods and other related topics. Some text was copied directly from the source when it was appropriately concise. The document is broken into 9 categories: Forest Pollinator Biology, Phenology, Habitat; Forest Pollinators and Woody Material; Forest Management and Pollinators; Forest Pollinators and Agriculture; Forest Pollinators and Habitat Fragments; Forest Pollinators and Adjacent Land Use; Forest Pollinators and Invasive Plants; Pollinator Research Methodology; and Miscellaneous, for papers that do not fit a specific category but that contain relevant information. Forest Pollinator Biology, Phenology, Habitat 1. Adamson, Nancy Lee, et al. “Pollinator Plants: Great Lakes Region,” Xerces Society for Invertebrate Conservation, 2017. Includes a list of plants beneficial to pollinators native to the Great Lakes region, which includes land in Ontario, Minnesota, Wisconsin, Ohio, Michigan, Pennsylvania, and New York. 2. Adamson, Nancy Lee, et al. “Pollinator Plants: Northeast Region,” Xerces Society for Invertebrate Conservation, 2015. Includes a list of plants beneficial to pollinators native to the Northeast Region, which encompasses southern Quebec, New Brunswick, Nova Scotia, the New England states, and eastern New York. 3. Black, Scott Hoffman, et al. “Pollinators in Natural Areas: A Primer on Habitat Management,” Xerces Society for Invertebrate Conservation. By aiding in wildland food production, helping with nutrient cycling, and as direct prey, pollinators are important in wildlife food webs. Summerville and Crist (2002) found that forest moths play important functional roles as selective herbivores, pollinators, detritivores, and prey for migratory songbirds. Belfrage et al. (2005) demonstrated that butterfly diversity was a good predictor of bird abundance and diversity, apparently due to a shared requirement for a complex plant community. -

626 Votants: 263 Blancs Et Nuls: 3 Suffrage Exprimes: 260
RESULTAT LEGISLATIVES 27 MAI 2019 District: IFANADIANA Commune: AMBOHIMANGA SUD Code Bureau: 350101010101 AMBOHIMANGA SUD EPP AMBOHIMANGA SUD INSCRITS: 626 VOTANTS: 263 BLANCS ET NULS: 3 SUFFRAGE EXPRIMES: 260 N° Partie Voix Poucentage 1 IRD 123 47,31% 2 MAFI 6 2,31% 3 TIM 128 49,23% 4 INDEPENDANT RABEMIAFARA MANARINTSOA JOSÉ 3 1,15% Total des voix 260 RESULTAT LEGISLATIVES 27 MAI 2019 District: IFANADIANA Commune: AMBOHIMANGA SUD Code Bureau: 350101020101 SAHAMARANA EPP SAHAMARANA INSCRITS: 359 VOTANTS: 86 BLANCS ET NULS: 4 SUFFRAGE EXPRIMES: 82 N° Partie Voix Poucentage 1 IRD 51 62,20% 2 MAFI 3 3,66% 3 TIM 27 32,93% 4 INDEPENDANT RABEMIAFARA MANARINTSOA JOSÉ 1 1,22% Total des voix 82 RESULTAT LEGISLATIVES 27 MAI 2019 District: IFANADIANA Commune: AMBOHIMANGA SUD Code Bureau: 350101030101 MANDROALINA EPP MANDROALINA INSCRITS: 302 VOTANTS: 103 BLANCS ET NULS: 2 SUFFRAGE EXPRIMES: 101 N° Partie Voix Poucentage 1 IRD 71 70,30% 2 MAFI 1 0,99% 3 TIM 22 21,78% 4 INDEPENDANT RABEMIAFARA MANARINTSOA JOSÉ 7 6,93% Total des voix 101 RESULTAT LEGISLATIVES 27 MAI 2019 District: IFANADIANA Commune: AMBOHIMANGA SUD Code Bureau: 350101040101 AMBALAHERANA EPP AMBALAHERANA INSCRITS: 176 VOTANTS: 118 BLANCS ET NULS: 0 SUFFRAGE EXPRIMES: 118 N° Partie Voix Poucentage 1 IRD 87 73,73% 2 MAFI 1 0,85% 3 TIM 30 25,42% 4 INDEPENDANT RABEMIAFARA MANARINTSOA JOSÉ 0 0,00% Total des voix 118 RESULTAT LEGISLATIVES 27 MAI 2019 District: IFANADIANA Commune: AMBOHIMANGA SUD Code Bureau: 350101050101 AMBALAVARY EPP AMBALAVARY INSCRITS: 166 VOTANTS: 120 BLANCS ET NULS: -

Does Flowering Synchrony Contribute to the Sustainment of Dry Grassland
Flora 222 (2016) 96–103 Contents lists available at ScienceDirect Flora j ournal homepage: www.elsevier.com/locate/flora Does flowering synchrony contribute to the sustainment of dry grassland biodiversity? a,∗ a a b Edy Fantinato , Silvia Del Vecchio , Antonio Slaviero , Luisa Conti , b a Alicia Teresa Rosario Acosta , Gabriella Buffa a Department of Environmental Sciences, Informatics and Statistics, Ca’ Foscari University of Venice, Via Torino 155, 30172 Venice, Italy b Department of Sciences, Roma Tre University, Viale G. Marconi 446, 00146 Rome, Italy a r t i c l e i n f o a b s t r a c t Article history: Phenological relationships among entomophilous species for pollination may play an important role in Received 20 January 2016 structuring natural plant communities. Received in revised form 1 April 2016 The main aim of this work was to test whether in dry grassland communities there is a non-random Accepted 7 April 2016 flowering pattern and if the pattern influences the species richness, and the richness of subordinate and Edited by Fei-Hai Yu common species. Available online 11 April 2016 Field sampling was carried out in temperate dry grasslands in NE Italy. Species composition and the flowering phenology were monitored in 45 2 m × 2 m plots randomly placed over dry grasslands. Keywords: To quantify the degree to which insect-pollinated species overlap in their flowering time we developed Co-flowering index a “co-flowering index” (CF-index). The significance of the observed flowering pattern was tested using a Generalists and specialists species Pollinator sharing null model. -

Pollen Limitation and Flower Abortion in a Wind-Pollinated, Masting Tree
Notes Ecology, 96(2), 2015, pp. 587–593 Ó 2015 by the Ecological Society of America Pollen limitation and flower abortion in a wind-pollinated, masting tree 1,2,5 1,3 4 1 IAN S. PEARSE, WALTER D. KOENIG, KYLE A. FUNK, AND MARIO B. PESENDORFER 1Cornell Lab of Ornithology, 159 Sapsucker Woods Road, Ithaca, New York 14850 USA 2Illinois Natural History Survey, 1816 S. Oaks Street, Champaign, Illinois 61820 USA 3Department of Neurobiology and Behavior, Cornell University, Ithaca, New York 14853 USA 4School of Biological Sciences, University of Nebraska, Lincoln, Nebraska 68588 USA Abstract. Pollen limitation is a key assumption of theories that explain mast seeding, which is common among wind-pollinated and woody plants. In particular, the pollen coupling hypothesis and pollination Moran effect hypothesis assume pollen limitation as a factor that synchronizes seed crops across individuals. The existence of pollen limitation has not, however, been unambiguously demonstrated in wind-pollinated, masting trees. We conducted a two-year pollen supplementation experiment on a masting oak species, Quercus lobata. Supplemental pollen increased acorn set in one year but not in the other, supporting the importance of pollen coupling and pollination Moran effect models of mast seeding. We also tracked the fate of female flowers over five years and found that the vast majority of flowers were aborted for reasons unrelated to pollination, even in the presence of excess pollen. Pollen limitation can reduce annual seed set in a wind-pollinated tree, but factors other than pollen limitation cause the majority of flower abortion. Key words: anemophily; flower abortion; perennial plants; pollen; seed set; wind pollination. -

Behavior of Pollinators That Share Two Co-Flowering Wetland Plant Species" (2015)
The University of Akron IdeaExchange@UAkron The Dr. Gary B. and Pamela S. Williams Honors Honors Research Projects College Spring 2015 Behavior of Pollinators That Share Two Co- Flowering Wetland Plant Species Joshua R. Morris University of Akron Main Campus, [email protected] Please take a moment to share how this work helps you through this survey. Your feedback will be important as we plan further development of our repository. Follow this and additional works at: http://ideaexchange.uakron.edu/honors_research_projects Part of the Animal Studies Commons, Biology Commons, and the Population Biology Commons Recommended Citation Morris, Joshua R., "Behavior of Pollinators That Share Two Co-Flowering Wetland Plant Species" (2015). Honors Research Projects. 56. http://ideaexchange.uakron.edu/honors_research_projects/56 This Honors Research Project is brought to you for free and open access by The Dr. Gary B. and Pamela S. Williams Honors College at IdeaExchange@UAkron, the institutional repository of The nivU ersity of Akron in Akron, Ohio, USA. It has been accepted for inclusion in Honors Research Projects by an authorized administrator of IdeaExchange@UAkron. For more information, please contact [email protected], [email protected]. Morris 1 Behavior of Pollinators That Share Two Co-Flowering Wetland Plant Species Joshua Morris Department of Biology Honors Research Project Morris 2 Behavior of Pollinators that share Two Co-Flowering Wetland Plant Species Abstract: Intermixed, co-flowering plant species often attract the same pollinators and may therefore compete for pollinator visits. Mimulus ringens and Verbena hastata are sympatric wetland plants that flower in synchrony and share many pollinators, the most common being bumblebees. -

Download Final Performance Evaluation of the Fararano
Final Performance Evaluation of the Fararano Development Food Security Activity in Madagascar March 2020 |Volume I IMPEL | Implementer-Led Evaluation & Learning Associate Award ABOUT IMPEL The Implementer-led Evaluation & Learning Associate Award works to improve the design and implementation of Food for Peace (FFP) funded development food security activities (DFSAs) through implementer-led evaluations and knowledge sharing. Funded by the USAID Office of Food for Peace (FFP), the Implementer-led Evaluation & Learning Associate Award will gather information and knowledge in order to measure performance of DFSAs, strengthen accountability, and improve guidance and policy. This information will help the food security community of practice and USAID to design projects and modify existing projects in ways that bolster performance, efficiency and effectiveness. The Implementer-led Evaluation & Learning Associate Award is a two-year activity (2019-2021) implemented by Save the Children (lead), TANGO International, and Tulane University in Haiti, the Democratic Republic of Congo, Madagascar, Malawi, Nepal, and Zimbabwe. RECOMMENDED CITATION IMPEL. (2020). Final Performance Evaluation of the Fararano Development Food Security Activity in Madagascar (Vol. 1). Washington, DC: The Implementer-Led Evaluation & Learning Associate Award PHOTO CREDITS Three-year-old child, at home in Mangily village (Toliara II District), after recovering from moderate acute malnutrition thanks to support from the Fararano Project. Photo by Heidi Yanulis for CRS. DISCLAIMER This report is made possible by the generous support of the American people through the United States Agency for International Development (USAID). The contents are the responsibility of the Implementer-Led Evaluation & Learning (IMPEL) award and do not necessarily reflect the views of USAID or the United States Government. -

Le Developpement Economique De La Region Vatovavy Fitovinany
UNIVERSITE D’ANTANANARIVO Année Universitaire : 2006-2007 Faculté de Droit, d’Economie, de Second Cycle – Promotion Sortante Gestion et de Sociologie Option : DEVELOPPEMENT DEPARTEMENT ECONOMIE « Promotion ANDRAINA » Mémoire de fin de Cycle LE DEVELOPPEMENT ECONOMIQUE DE LA REGION VATOVAVY FITOVINANY Encadré par : Monsieur Gédéon RAJAONSON Présenté par : MANIRISOA RAZAFIMARINTSARA Firmin Date de soutenance : 14 Décembre 2007 REMERCIEMENTS Pour commencer, je tiens à exprimer toute ma reconnaissance à tous ceux qui ont contribué, de près ou de loin, à ma formation et à la réalisation de ce Grand Mémoire de fin d’études en Economie. J’adresse donc tout particulièrement mes vifs remerciement à : • DIEU TOUT PUISSANT • Mon encadreur Monsieur Gédéon RAJAONSON ; • Tous les enseignants et les Personnels administratifs du Département Economie de la Faculté DEGS de l’Université d’Antananarivo ; • Monsieur Le Chef de Région de Vatovavy Fitovinany et ses équipes • Monsieur le Directeur Régional des Travaux Publics de Vatovavy Fitovinany • Ma famille pour leurs soutiens permanents. Veuillez accepter le témoignage de ma profonde gratitude. LISTE DES ABREVIATIONS ANGAP : Agence Nationale de la Gestion des Aires Protégés CEG : Collège d’Enseignement Général CHD 1 : Centre Hospitalier de District Niveau 1 CHD 2 : Centre Hospitalier de District Niveau 2 CISCO : Circonscription Scolaire CSB 1 : Centre de Santé de Base Niveau 1 CSB 2 : Centre de Santé de Base Niveau 2 DRDR : Direction Régionale du Développement Rural EPP : Ecole Primaire Public FCE : Fianarantsoa Côte Est FER : Fonds d’Entretien Routier FTM : Foibe Toantsritanin’i Madagasikara GU : Guichet Unique HIMO : Haute Intensité de Main d’œuvre INSTAT : Institut National de la Statistique M.A.E.P.