Gene Expression Part V
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
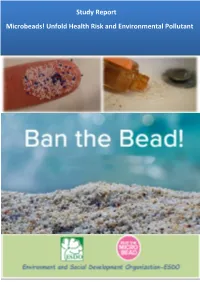
Study Report Microbeads! Unfold Health Risk and Environmental Pollutant
Study Report Microbeads! Unfold Health Risk and Environmental Pollutant Study Report: Microbeads! Unfold Health Risk and Environmental Pollutant- Copyright © ESDO,2016 BD | Page 2 Study Report: Microbeads! Unfold Health Risk and Environmental Pollutant- Copyright © ESDO,2016 BD | Page 3 Study Report Microbeads! Unfold Health Risk and Environmental Pollutant Advisor Syed Marghub Murshed, Prof. Dr. Abu Jafar Mahmud, Prof. Dr. Md. Abul Hashem, Dr. Shahriar Hossain Study Advisory Team Syed Marghub Murshed, Dr. Shahriar Hossain . Team Leader Siddika Sultana Editorial Team Farida Shahnaz, Sayda Mehrabin Shejuti. Research Associate Nishat Ferdousi Support Team Md. Ali Hossain, Md. Mamun Ul Hasan, Sayda Mehrabin Shejuti , Md. Asif Iqbal. Study Period January2015- October, 2016. Report Published October 15, 2016 Copyright © Environment and Social Development Organization-ESDO, 2016, all rights reserved Disclaimer "While this study and publication have been produced under the organization core funding and direct supervision of research and management team of ESDO, the contents of the publication are the sole responsibility of ESDO. In addition, ESDO has core focussed on plastic pollution and marine littering with the overall objectives of raising public awareness, strengthing capaciity of institutional reserch, policy advocacy to reduce and eliminate the microbeads/microplastics to protect public health and environment in Bangladesh." Contact Address: House # 8/1, Level # 5, Block # C, Lalmatia, Dhaka-1207, Bangladesh, Phone: 880-2-912-2729 Fax: -

SIDR: Simultaneous Isolation and Parallel Sequencing of Genomic DNA and Total RNA from Single Cells
Downloaded from genome.cshlp.org on October 3, 2021 - Published by Cold Spring Harbor Laboratory Press 1 SIDR: simultaneous isolation and parallel sequencing of genomic DNA and 2 total RNA from single cells 3 4 Kyung Yeon Han1,9, Kyu-Tae Kim1,9, Je-Gun Joung1,9, Dae-Soon Son1, Yeon Jeong Kim1, Areum Jo1, 5 Hyo-Jeong Jeon1, Hui-Sung Moon1, Chang Eun Yoo1, Woosung Chung1,2, Hye Hyeon Eum1,3, 6 Sangmin Kim4, Hong Kwan Kim5, Jeong Eon Lee2,6, Myung-Ju Ahn7, Hae-Ock Lee1,8, Donghyun 7 Park1,**, Woong-Yang Park1,2,8,* 8 1 Samsung Genome Institute, Samsung Medical Center, Seoul 06351, South Korea 9 2 Department of Health Sciences and Technology, SAIHST, Sungkyunkwan University, Seoul 06351, 10 South Korea 11 3 Department of Biomedical Sciences, College of Medicine, Seoul National University, Seoul 03080, 12 South Korea 13 4 Department of Breast Cancer Center, Samsung Medical Center, Seoul 06351, South Korea 14 5 Department of Thoracic and Cardiovascular Surgery, Samsung Medical Center, Seoul 06351, South 15 Korea 16 6 Department of Surgery, Samsung Medical Center, Sungkyunkwan University School of Medicine, 17 Seoul, 06351, South Korea 18 7 Division of Hematology-Oncology, Department of Medicine, Samsung Medical Center, 19 Sungkyunkwan University School of Medicine, Seoul 06351, South Korea 20 8 Department of Molecular Cell Biology, Sungkyunkwan University School of Medicine, Suwon 16419, 21 South Korea 22 23 * Corresponding author. Tel.: +82 2 2148 9810; fax: +82 2 2148 9819. 24 ** Corresponding author. Tel.: +82 2 3410 2954; fax: +82 2 2148 9819. 25 E-mail addresses: [email protected] (D. -

Deciphering the Effect of Microbead Size Distribution on the Kinetics of Heterogeneous Biocatalysts Through Single-Particle Anal
catalysts Article Deciphering the Effect of Microbead Size Distribution on the Kinetics of Heterogeneous Biocatalysts through Single-Particle Analysis Based on Fluorescence Microscopy Emilio Muñoz-Morales 1, Susana Velasco-Lozano 1, Ana I. Benítez-Mateos 2, María J. Marín 3, Pedro Ramos-Cabrer 4,5 and Fernando López-Gallego 1,2,5,6,* 1 Heterogeneous Biocatalysis Laboratory, Instituto de Síntesis Química y Catálisis Homogénea (iSQCH), CSIC-Universidad de Zaragoza, C/Pedro Cerbuna 12, 50009 Zaragoza, Spain; [email protected] (E.M.-M.); [email protected] (S.V.-L.) 2 Heterogeneous Biocatalysis Laboratory, CICbiomaGUNE, Edificio Empresarial “C”, Paseo de Miramón, 182, 20014 Donostia-San Sebastián, Spain; [email protected] 3 Servicio General de Apoyo a la Investigación—SAI, Servicio de Microscopía Electrónica de Sistemas Biológicos, Universidad de Zaragoza, C/Pedro Cerbuna 12, 50009 Zaragoza, Spain; [email protected] 4 Magnetic Resonance Imaging Lab, CICbiomaGUNE, Edificio Empresarial “C”, Paseo de Miramón, 182, 20014 Donostia-San Sebastián, Spain; [email protected] 5 IKERBASQUE, Basque Foundation for Science, Maria Diaz de Haro 3, 48013 Bilbao, Spain 6 ARAID, Aragon foundation for Science, 50009 Zaragoza, Spain * Correspondence: fl[email protected] Received: 2 October 2019; Accepted: 24 October 2019; Published: 28 October 2019 Abstract: Understanding the functionality of immobilized enzymes with spatiotemporal resolution and under operando conditions is an unmet need in applied biocatalysis, as well as priceless information to guide the optimization of heterogeneous biocatalysts for industrial purposes. Unfortunately, enzyme immobilization still relies on trial-and-error approximations that prevail over rational designs. Hence, a modern fabrication process to achieve efficient and robust heterogeneous biocatalysts demands comprehensive characterization techniques to track and understand the immobilization process at the protein–material interface. -

A Technique That Uses Microscale Beads to Determine Cell Biomechanics…
A technique that uses microscale beads to determine cell biomechanics… 1 Professor John Ugelstad (Univ. of Trondheim, Norway) greatest scientific achievement was the preparation of large monodisperse polymer beads in 1977 (today known as DynaBeads). Previously, this was only achievable by NASA in the weightlessness of space. The basic idea for this invention was a two-step swelling procedure. Small "activated" polymer particles are dispersed in water, and these will adsorb slightly water-soluble organic compounds and reach a volume more than 1000 times their own volume. Further developments resulted in preparation of monodisperse paramagnetic beads in 1981. These may be prepared by dispersing porous beads covered with oxidative groups at the exterior and interior surfaces in an aqueous solution of Fe 2+ containing salts. The ions react with the oxidative groups and form fine grains of magnetic oxides throughout the beads, resulting in supermagnetic beads, i.e. the beads are only magnetic in a magnetic field. Many applications of these beads lie within immunology, cellular biology, microbiology, molecular biology, medical diagnostics and DNA technology 2 Two different approaches for cell biomechanics measurements have relied upon the colloidal technology of microparticles . The first is a force application system that uses magnetic fields to pull on the ferrous microbead. The second uses the beads as markers of the local brownian motion within a cell. These particles are injected inside the cell and their positions over a set time frame are tracked to determine their degree of freedom and mobility. From the movement of the particles, the elasticity and viscosity of the cell can be approximated. -

Fast and Easy Single-Molecule Pulldown Assay Based on Agarose Microbeads
bioRxiv preprint doi: https://doi.org/10.1101/2020.09.20.305177; this version posted September 20, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Fast and easy single-molecule pulldown assay based on agarose microbeads Qirui Zhao1, Yusheng Shen2, Xiaofen Li2, Fang Tian3, Xiaojie Yu1, Levent Yobas2,4, Hyokeun Park1,3, Pingbo Huang#1,2,5,6,7 Division of Life Science1, Department of Chemical and Biological Engineering2, Department of Physics3, Department of Electronic and Computer Engineering4, State Key Laboratory of Molecular Neuroscience5, HKUST Shenzhen Research Institute6, and Hong Kong Branch of Guangdong Southern Marine Science and Engineering Laboratory (Guangzhou)7, Hong Kong University of Science and Technology, Hong Kong, China. # To whom correspondence should be addressed: [email protected] SUMMARY The recently developed single-molecule pulldown (SiMPull) assay by Jain and colleagues is a highly innovative technique but its wide application is hindered by the high technical barrier and time consumption. We report an innovative, agarose microbead-based approach for SiMPull. We used commercially available, pre-surface- functionalized agarose microbeads to capture the protein of interest together with its binding partners specifically from cell extracts and observed these interactions under a microscope at the single-molecule level. Relative to the original method, microbead- based SiMPull is considerably faster, easier to use, and more reproducible and yet provides similar sensitivity and signal-to-noise ratio; specifically, with the new method, sample-preparation time is substantially decreased (from ~10 to ~3 h). -

DNA Detection Method Based on the Microbead Velocity Under Traveling Wave Dielectrophoresis
DNA Detection Method based on the Microbead Velocity under Traveling Wave Dielectrophoresis Zhenhao Ding1, Michihiko Nakano2 and Junya Suehiro2 1Graduate School of Information Science and Electrical Engineering, Kyushu University, 744 Motooka, Fukuoka, Japan 2Faculty of Information Science and Electrical Engineering, Kyushu University, 744 Motooka, Fukuoka, Japan Keywords: DNA Detection, Traveling Wave Dielectrophoresis, Microbeads. Abstract: Polymerase chain reaction (PCR) is a highly specific and sensitive detection method for bacterial and viral infections by amplifying the specific regions of DNA or RNA via enzymatic reaction. The authors have developed a rapid DNA detection method based on the dielectrophoresis (DEP) characteristic of DNA labeled microbeads for the rapid detection of the DNA that amplified by PCR. This method is based on the change of the Clausius-Mossotti (CM) factor K of DEP when DNA is attached onto microbeads. In former studies, we developed a new DNA detection method based on the change of the real part of K (Re[K]). However, this method requires a large amount of DNA attaching to a microbead to alter the microbead DEP behaviour for DNA detection. In this study, we focus on the imaginary part of K (Im[K]), which theoretically will change more dramatically than Re[K] when DNA is attached onto the microbeads. Since the traveling wave dielectrophoresis behaviour is based on the Im[K], we propose a new method for DNA detection based on the traveling wave dielectrophoresis (twDEP) of microbeads. 1 INTRODUCTION Mossotti (CM) factor K of DEP. When enough amount of DNA is attached to the microbeads, the There are various diseases caused by bacterial or real part of K (Re[K]) will change from negative to viral infections. -

Microbead-Based Raman/Surface Enhanced Raman
Microbead-based Raman/Surface Enhanced Raman Scattering Immunoassays for Multiplex Detection A THESIS SUBMITTED BY Lijuan Wei FOR THE DEGREE OF DOCTOR OF PHILOSOPHY School of Chemical Engineering The University of Adelaide, Australia October 2013 I dedicate this whole thesis to my beloved husband, for his support, encouragement and love. Declaration DECLARATION I certify that this work contains no material which has been accepted for the award of any other degree or diploma in any university or other tertiary institution and, to the best of my knowledge and belief, contains no material previously published or written by another person, except where due reference has been made in the text. In addition, I certify that no part of this work will, in the future, be used in a submission for any other degree or diploma in any university or other tertiary institution without the prior approval of the University of Adelaide and where applicable, any partner institution responsible for the joint-award of this degree. I give consent to this copy of my thesis when deposited in the University Library, being made available for loan and photocopying, subject to the provisions of the Copyright Act 1968. The author acknowledges that copyright of published works contained within this thesis resides with the copyright holder(s) of those works. I also give permission for the digital version of my thesis to be made available on the web, via the University’s digital research repository, the Library catalogue and also through web search engines, unless permission has been granted by the University to restrict access for a period of time. -

Sequence-Specific Detection of DNA Strands Using a Solid-State Nanopore Assisted by Microbeads
micromachines Article Sequence-Specific Detection of DNA Strands Using a Solid-State Nanopore Assisted by Microbeads Yin Zhang *, Zengdao Gu, Jiabin Zhao, Liying Shao and Yajing Kan Jiangsu Key Laboratory for Design and Manufacture of Micro-Nano Biomedical Instruments, School of Mechanical Engineering, Southeast University, Nanjing 211189, China; [email protected] (Z.G.); [email protected] (J.Z.); [email protected] (L.S.); [email protected] (Y.K.) * Correspondence: [email protected] Received: 19 November 2020; Accepted: 10 December 2020; Published: 11 December 2020 Abstract: Simple, rapid, and low-cost detection of DNA with specific sequence is crucial for molecular diagnosis and therapy applications. In this research, the target DNA molecules are bonded to the streptavidin-coated microbeads, after hybridizing with biotinylated probes. A nanopore with a diameter significantly smaller than the microbeads is used to detect DNA molecules through the ionic pulse signals. Because the DNA molecules attached on the microbead should dissociate from the beads before completely passing through the pore, the signal duration time for the target DNA is two orders of magnitude longer than free DNA. Moreover, the high local concentration of target DNA molecules on the surface of microbeads leads to multiple DNA molecules translocating through the pore simultaneously, which generates pulse signals with amplitude much larger than single free DNA translocation events. Therefore, the DNA molecules with specific sequence can be easily identified by a nanopore sensor assisted by microbeads according to the ionic pulse signals. Keywords: nanopore; microbead; DNA detection 1. Introduction The sequence-specific detection of DNA or RNA targets plays an important role in molecular diagnosis and therapy applications [1–4]. -

Spectral-Optical-Tweezer-Assisted Fluorescence Multiplexing System
Biosensors and Bioelectronics 129 (2019) 107–117 Contents lists available at ScienceDirect Biosensors and Bioelectronics journal homepage: www.elsevier.com/locate/bios Spectral-optical-tweezer-assisted fluorescence multiplexing system for QDs- T encoded bead-array bioassay ⁎ Qinghua Hea,b,1, Xuejing Chena,b,1, Yonghong Hea,b, Tian Guana,c, , Guangxia Fenga,c, ⁎⁎ Bangrong Lua, Bei Wanga, Xuesi Zhoua, Liangshan Hud, Donglin Caod, a Shenzhen Key Laboratory for Minimal Invasive Medical Technologies, Institute of Optical Imaging and Sensing, Graduate School at Shenzhen, Tsinghua University, Shenzhen 518055, China b Department of Physics, Tsinghua University, Beijing 100084, China c School of Medicine, Tsinghua University, Beijing 100084, China d Department of Laboratory Medicine, Guangdong Second Provincial General Hospital, Guangzhou 510317, China ARTICLE INFO ABSTRACT Keywords: As an efficient tool in the multiplexed detection of biomolecules, bead-array could achieve separation-free de- Optical tweezers tection to multiple targets, making it suitable to analyze valuable and scarce samples like antigen and antibody QDs encoding from living organism. Herein, we propose a spectral-optical-tweezer-assisted fluorescence multiplexing system to Bead-array analyze biomolecule-conjugated bead-array. Using optical tweezer, we trapped and locked beads at the focus to Multiplexed biodetection accept stimulation, offering a stable and optimized analysis condition. Moving the system focus and scanning the sample slide, we achieved emissions collection to QDs-encoded bead-array after the multiplexed detection. The emission spectra of fluorescence were collected and recorded by the spectrometer. By recognizing locations of decoding peaks and counting the intensities of label signals of emission spectra, we achieved qualitative and quantitative detection to targets. -

Notch Ligand Functionalized Microbead System for T Cell Differentiation
Notch ligand functionalized microbead system for T cell differentiation. Jia Wen Jessica Chang University of Texas at Austin 2007 Supervising Professor: Krishnendu Roy, Ph.D. _______________________________ Signature Date Honors/Dean’s Scholar Advisor: Shelley Payne, Ph.D. _______________________________ Signature Date Pg. 1 Table of Contents Abstract ………………………………………... Pg. 4 Introduction ………………………………………... Pg. 6 Materials and Methods ………………………………………... Pg. 17 Results ………………………………………... Pg. 20 Discussion ………………………………………... Pg. 28 References ………………………………………... Pg. 34 Pg. 2 ABSTRACT Hematopoietic stem cells (HSC) are cells that can self-replicate and give rise to a wide array of different cell types through the process of differentiation. HSC are found in blood and bone marrow, and they are responsible for the daily production of blood and immune cells essential to humans’ survival. HSCs’ multipotency makes it a powerful candidate for new, revolutionary treatments for diseases. However, in order to harness its multpotency, we must first understand what instructs HSC to become very specific cell types. In particular, we are interested in the role of the Notch signaling pathway in HSC differentiation into T-cells, white blood cells that regulate the immune system and destroy tumor and virus-infected cells. The Notch signaling pathway been shown to be necessary and sufficient for directing HSC differentiation into T-cells. Specifically we limit our studies to two proteins in the pathway: Notch1 receptor on the signal receiving cell’s membrane and delta-like ligand 4, a protein that activates Notch1. However, scalable and efficient biomaterial-based systems for this differentiation process have yet to be reported. Such a system would allow unprecedented temporal and ligand density control, prerequisites for the ultimate goal of mass producing T-cells “on demand” for therapeutic treatments for immunodeficient or cancer patients. -

Realtime Monitoring and Control of Soluble Signaling Factors Enables
COMMUNICATION TO THE EDITOR Real-Time Monitoring and Control of Soluble Signaling Factors Enables Enhanced Progenitor Cell Outputs from Human Cord Blood Stem Cell Cultures Elizabeth Csaszar,1,2 Kun Chen,1 Julia Caldwell,1 Warren Chan,1,2,3,4,5 Peter W. Zandstra1,2,3,6,7 1 Institute for Biomaterials and Biomedical Engineering, University of Toronto, Toronto, Ontario M5S3E1, Canada; telephone: þ416-978-8888; fax: þ416-978-2666; e-mail: [email protected] 2 Department of Chemical Engineering and Applied Chemistry, University of Toronto, Toronto, Ontario M5S3E1, Canada 3 Terrence Donnelly Centre for Cellular and Biomolecular Research, University of Toronto, Toronto, Ontario M5S3E1, Canada 4 Department of Chemistry, University of Toronto, Toronto, Ontario 5 Department of Materials Science and Engineering, University of Toronto, Toronto, Ontario 6 Heart and Stroke/Richard Lewar Centre of Excellence, University of Toronto, Toronto, Ontario M5G1L7, Canada 7 McEwen Centre for Regenerative Medicine, University Health Network, Toronto Medical Discovery Tower, Toronto, Ontario M5G1L7, Canada umbilical cord blood cell expansion system. By maintaining b ABSTRACT: Monitoring and control of primary cell cultures the concentration of TGF- 1 below an upper threshold is challenging as they are heterogenous and dynamically throughout the culture, we demonstrate enhanced ex vivo complex systems. Feedback signaling proteins produced from expansion of hematopoietic progenitor cells at higher input off-target cell populations can accumulate, inhibiting the cell densities and over longer culture periods. This study production of the desired cell populations. Although culture demonstrates the potential of a fully automated and strategies have been developed to reduce feedback inhibition, integrated real-time control strategy in stem cell culture they are typically optimized for a narrow range of process systems, and provides a powerful strategy to achieve highly fi parameters and do not allow for a dynamically regulated regulated and intensi ed in vitro cell manufacturing systems. -

Fast-Degradable Microbeads Encapsulating Human Umbilical Cord Stem Cells in Alginate for Muscle Tissue Engineering
TISSUE ENGINEERING: Part A Volume 18, Numbers 21 and 22, 2012 ª Mary Ann Liebert, Inc. DOI: 10.1089/ten.tea.2011.0658 Fast-Degradable Microbeads Encapsulating Human Umbilical Cord Stem Cells in Alginate for Muscle Tissue Engineering Jun Liu, D.D.S., Ph.D.,1,2 Hongzhi Zhou, D.D.S., Ph.D.,1 Michael D. Weir, Ph.D.,1 Hockin H.K. Xu, Ph.D.,1,3–5 Qianming Chen, D.D.S., Ph.D.,2 and Carroll Ann Trotman, B.D.S., M.A., M.S.6,7 Human umbilical cord mesenchymal stem cells (hUCMSCs) are inexhaustible and can be obtained without an invasive surgery. To date, there has been no report on seeding hUCMSCs in three-dimensional scaffolds for muscle tissue engineering. The objectives of this study were to (1) investigate hUCMSC seeding in a scaffold for muscle engineering and (2) develop a novel construct consisting of hUCMSC-encapsulating and fast- degradable microbeads inside a hydrogel matrix. The rationale was that the hydrogel matrix would maintain the defect volume, while the microbeads would degrade to release the cells and concomitantly create mac- ropores in the matrix. hUCMSCs were encapsulated in alginate-fibrin microbeads, which were packed in an Arg-Gly-Asp (RGD)-modified alginate matrix (AM). This construct is referred to as hUCMSC-microbead-AM. The control consisted of the usual cell encapsulation in AM without microbeads (referred to as hUCMSC-AM). In the hUCMSC-AM construct, the hUCMSCs showed as round dots with no spreading at 1–14 days. In contrast, cells in the hUCMSC-microbead-AM construct had a healthy spreading and elongated morphology.