<I>Phytophthora</I> Taxa Associated with Cultivated <I>Agathosma</I
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Effect of Rutaceae Plant's Essential Oils and Leaf Extracts on Dermatophytic Fungal Cell Morphology
Effect of Rutaceae plant’s essential oils and leaf extracts on dermatophytic fungal cell morphology: a hope for the development of an effective antifungal from natural origin Submitted in fulfillment of the academic requirements for the degree of DOCTOR OF PHILOSOPHY By OLUFUNKE OMOWUMI FAJINMI Research Centre for Plant Growth and Development School of Life Sciences College of Agriculture, Engineering and Science University of KwaZulu-Natal, Pietermaritzburg May 2016 Pictures sourced from google A healthy, glowing, beautiful skin….the pride of every woman i . Table of Contents STUDENT DECLARATION ................................................................................................... v DECLARATION BY SUPERVISORS ....................................................................................... vi COLLEGE OF AGRICULTURE ENGINEERING & SCIENCE DECLARATION 1- PLAGIARISM ........ vii ACKNOWLEDGEMENTS .................................................................................................. viii COLLEGE OF AGRICULTURE ENGINEERING & SCIENCE DECLARATION 2- PUBLICATIONS ....... x LIST OF FIGURES .............................................................................................................. xi LIST OF TABLES ............................................................................................................... xii LIST OF ABBREVIATIONS ................................................................................................ xiv ABSTRACT ..................................................................................................................... -

Obdiplostemony: the Occurrence of a Transitional Stage Linking Robust Flower Configurations
Annals of Botany 117: 709–724, 2016 doi:10.1093/aob/mcw017, available online at www.aob.oxfordjournals.org VIEWPOINT: PART OF A SPECIAL ISSUE ON DEVELOPMENTAL ROBUSTNESS AND SPECIES DIVERSITY Obdiplostemony: the occurrence of a transitional stage linking robust flower configurations Louis Ronse De Craene1* and Kester Bull-Herenu~ 2,3,4 1Royal Botanic Garden Edinburgh, Edinburgh, UK, 2Departamento de Ecologıa, Pontificia Universidad Catolica de Chile, 3 4 Santiago, Chile, Escuela de Pedagogıa en Biologıa y Ciencias, Universidad Central de Chile and Fundacion Flores, Ministro Downloaded from https://academic.oup.com/aob/article/117/5/709/1742492 by guest on 24 December 2020 Carvajal 30, Santiago, Chile * For correspondence. E-mail [email protected] Received: 17 July 2015 Returned for revision: 1 September 2015 Accepted: 23 December 2015 Published electronically: 24 March 2016 Background and Aims Obdiplostemony has long been a controversial condition as it diverges from diploste- mony found among most core eudicot orders by the more external insertion of the alternisepalous stamens. In this paper we review the definition and occurrence of obdiplostemony, and analyse how the condition has impacted on floral diversification and species evolution. Key Results Obdiplostemony represents an amalgamation of at least five different floral developmental pathways, all of them leading to the external positioning of the alternisepalous stamen whorl within a two-whorled androe- cium. In secondary obdiplostemony the antesepalous stamens arise before the alternisepalous stamens. The position of alternisepalous stamens at maturity is more external due to subtle shifts of stamens linked to a weakening of the alternisepalous sector including stamen and petal (type I), alternisepalous stamens arising de facto externally of antesepalous stamens (type II) or alternisepalous stamens shifting outside due to the sterilization of antesepalous sta- mens (type III: Sapotaceae). -

Kirstenbosch NBG List of Plants That Provide Food for Honey Bees
Indigenous South African Plants that Provide Food for Honey Bees Honey bees feed on nectar (carbohydrates) and pollen (protein) from a wide variety of flowering plants. While the honey bee forages for nectar and pollen, it transfers pollen from one flower to another, providing the service of pollination, which allows the plant to reproduce. However, bees don’t pollinate all flowers that they visit. This list is based on observations of bees visiting flowers in Kirstenbosch National Botanical Garden, and on a variety of references, in particular the following: Plant of the Week articles on www.PlantZAfrica.com Johannsmeier, M.F. 2005. Beeplants of the South-Western Cape, Nectar and pollen sources of honeybees (revised and expanded). Plant Protection Research Institute Handbook No. 17. Agricultural Research Council, Plant Protection Research Institute, Pretoria, South Africa This list is primarily Western Cape, but does have application elsewhere. When planting, check with a local nursery for subspecies or varieties that occur locally to prevent inappropriate hybridisations with natural veld species in your vicinity. Annuals Gazania spp. Scabiosa columbaria Arctotis fastuosa Geranium drakensbergensis Scabiosa drakensbergensis Arctotis hirsuta Geranium incanum Scabiosa incisa Arctotis venusta Geranium multisectum Selago corymbosa Carpanthea pomeridiana Geranium sanguineum Selago canescens Ceratotheca triloba (& Helichrysum argyrophyllum Selago villicaulis ‘Purple Turtle’ carpenter bees) Helichrysum cymosum Senecio glastifolius Dimorphotheca -

Agathosma Betulina Herba
AGATHOSMA BETULINA HERBA Definition Agathosma Betulina Herba consists of the fresh or dried leaves and smaller stalks of Agathosma betulina (Berg.) Pillans (Rutaceae). Synonyms Barosma betulina Bartl. and Wendl. f. Hartogia betulina Berg. Vernacular names Figure 1a: Live plant boegoe, bergboegoe (A), buchu (San); round leaf buchu Description Macroscopical 1, 2 Evergreen, multi-stemmed, perennial resprouting woody shrub to 1m in height, with glabrous yellow to red-brown stems; leaves alternate to opposite, 14-25 × 6- Figure 1b: Dried leaf 14mm, broadly elliptic to nearly round (average length:breadth ratio 1.95), with rounded and recurved apex; glabrous with prominent main and subsidiary veins on abaxial surface; gland dotted on underside; margin serrate with an oil gland at the base of each serration; flowers (June-Nov) axillary, usually solitary, up to 20mm in diameter, white to pale purple-pink, borne on slender stalks ±7mm long. Figure 2: line drawing 1 Pillans, N. (1950). A revision of the genus Agathosma (Rutaceae). Journal of South African Botany 16: 55-117. 2 Spreeth, A.D. (1976). ʼn Hersiening van die Agathosma-species van kommersiële belang (A revision of the commercially important Agathosma species). Journal of South African Botany 42(2): 109-119. Microscopical 3 Crude drug Collected as required or available in the marketplace as bundles of leafy twigs with light yellow-green, highly aromatic foliage; texture soft when fresh, leathery when dry; occasional flowers may be present. BPC quality buchu leaf is freely available in pharmacies in South Africa and unstandardised leaf in supermarkets. Geographical distribution Sandy mountain slopes of the Western Cape Province, in the Calvinia, Cedarberg, Tulbagh, Ceres and Piketberg districts, at altitudes of 300-700m above sea level. -

A Phylogeny of the Rutaceae and a Biogeographic Study of Its Subfamily Aurantioideae
A phylogeny of the Rutaceae and a biogeographic study of its subfamily Aurantioideae Thomas Schwartz Supervisor Bernard Pfeil Degree project for Master of Science In Systematics and Biodiversity, Biology 60 hec Department of Plant and Environmental Science University of Gothenburg Abstract The Rutaceae classification is complex and has undergone several changes. In addition to morphological studies, phylogenetic inference using molecular data has also led to classification changes. Thus far only chloroplast data and ITS have been used, sometimes combined with morphology to infer the phylogeny. This study adds information from a low copy nuclear gene to test the existing phylogenetic hypothesis using a species tree framework. A biogeographic study was also performed on the Aurantioideae subfamily. A pilot study looked at the choice of genes, followed by testing and evaluation of several methods for extraction of Rutaceae DNA. Thereafter, a new method for efficient separation of alleles and paralogues was examined. The sequences obtained were analysed for recombination, positive selection and hybridisation. Trees for three loci (chloroplast, nuclear HYB and MDH) were made using MrBayes and BEAST, and a species tree was constructed with *BEAST. The *BEAST species tree is used as a template for a biogeographic study with the Lagrange geographic range likelihood analysis. A Bayesian biogeographic study is also performed using a Bayesian discrete biogeographical mode (an addition to BEAST). The results are then compared with previous studies, corroborating some and rejecting others. Sammanfattning Rutace-familjens struktur är komplex och föränderlig. Förutom morfologiska studier har även fylogenetiska studier använts för att få ordning i den. Hittills har man endast tittat på kloroplastgener och ribosom-DNA, i enstaka fall i kombination med morfologiska karaktärer. -

Buchu Oil Its Characteristic Blackcurrant What's in a Name? Smell and Flavour but the Profiles Differ Between Species
give buchu oil its characteristic blackcurrant what'S IN A NAME? smell and flavour but the profiles differ between species. The possibly toxic pulegone Botanical names: The three buchu species harvested commercially from the wild are Agathosma betulina, A. crenulata is found in high concentrations (50%) in and A. serratifolia. Agathosma is derived from the Greek agathos (good) and osme (scent), referring to the fragrance of A. crenulata, giving its oil a sharp smell. Pulegone its leaves. is only minimally present in A. betulina (less Family name: Rutaceae, the citrus family, many of whose members have leaves with conspicuous oil glands and a sharp than 3%), the favoured choice for the local lemon-like scent when crushed. and export food, medicinal and bottled water Common names: A. betulina: Buchu (Khoi), Round-leaf Buchu (Eng.), iBuchu (Xhosa), Rondeblaarboegoe, Bergboegoe flavour markets. Pharmacological analysis of (Afr.). A crenulata: Oval-leaf Buchu (Eng.), Anysboegoe (Afr.). A serratifolia: Long-leaf Buchu (Eng.), Kloofboegoe (Afr.). the essential oil and extracts show weak anti- European herbal books often still refer to buchu species under the old generic synonym Barosma. They distinguish between Short Buchu, or ‘shorts’ (A. betulina), Oval Buchu, or ‘ovals’ or ‘shortbroads’ (A. crenulata) and Long Buchu, or microbial, anti-oxidant, anti-inflammatory and ‘longs’ (A. serratifolia). anti-spasmodic activity. Buchu is also popularly available as a tonic and digestive tea, and a brandy tincture developed by early Dutch colonists, ‘boegoebrandewyn’ is still taken for An easy guide to growing buchu many conditions. Commercial exploitation ENVIRONMENT Since demand for dried buchu leaves and ACTIVITY REQUIRED FOR TREATMENT TIME REMARKS SUCCESS the steam-distilled oil extract far outstrips production, sustainable commercialization Harvest Fruit or seed Seed can be stored in a brown Seed ripens The seeds collected from fully ripe seed capsules develop paper bag in a ventilated area. -

Planta Medica
www.thieme-connect.de/ejournals | www.thieme.de/fz/plantamedica Planta Medica August 2010 · Page 1163 – 1374 · Volume 76 12 · 2010 1163 Editorial 1177 Special Session: Opportunities and challenges in the exploitation of biodiversity – Complying with the principles of the convention on biological diversity th 7 Tannin Conference 1178 Short Lectures 1164 Lectures 1193 Posters 1165 Short Lectures 1193 Aphrodisiaca from plants 1193 Authentication of plants and drugs/DNA-Barcoding/ th 58 International Congress and Annual Meeting of PCR profiling the Society for Medicinal Plant and Natural Product Research 1197 Biodiversity 1167 Lectures 1208 Biopiracy and bioprospecting 1169 WS I: Workshops for Young Researchers 1169 Cellular and molecular mechanisms of action of natural 1208 Enzyme inhibitors from plants products and medicinal plants 1214 Fertility management by natural products 1171 WS II: Workshops for Young Researchers 1214 Indigenous knowledge of traditional medicine and 1171 Lead finding from Nature – Pitfalls and challenges of evidence based herbal medicine classical, computational and hyphenated approaches 1230 Miscellaneous 1173 WS III: Permanent Committee on Regulatory Affairs of Herbal Medicinal Products 1292 Natural products for the treatment of infectious diseases 1173 The importance of a risk-benefit analysis for the marketing authorization and/or registration of (tradi- 1323 New analytical methods tional) herbal medicinal products (HMPs) 1337 New Targets for herbal medicines 1174 WS IV: Permanent Committee on Biological and -
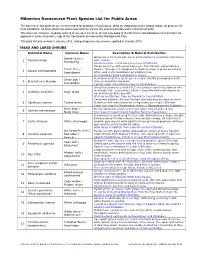
Plant List Are Recommended for Planting in Royal Ascot
1 Milnerton Racecourse Plant Species List for Public Areas The species on this plant list are recommended for planting in Royal Ascot. Most are indigenous to the fynbos region; all grow well in local conditions. All these plants are water-wise and can survive dry summer periods with a minimum of water. This list is not exclusive, anybody wishing to use species not on the list may apply to the Environmental Management Committee for approval in terms of section 2.6(g) of the Operational Environmental Management Plan. This plant list was revised in January 2012, changed species names were updated in January 2013. TREES AND LARGE SHRUBS Botanical Name Common Name Description & Natural Distribution Grows up to 15 m tall with sweet scented flowers in summer and silvery Sweet thorn / 1 Acacia karroo white thorns. Soetdoring Southern Africa – from low-lying areas to highveld. Fat growing tree with yellow bark up to 15m tall with yellow ball-like Fever tree / flowers. Athough not indigenous to Western Cape, it grows well and is 2 Acacia xanthophloea Koorsboom often used in city landscapes as a feature tree. From KwaZulu Natal norhtwards to Kenya. Silver oak / A shrub or small tree up to 10m in height. Usually developing a multi- 3 Brachylaena discolor stem or branching low down. Kusvaalbos Coastal region from Eastern Cape to Mozambique. Shrub that grows to a height of 5 m bearing cream to lilac flowers with an orange ‘eye’. In summer/ autumn. Sage-like dark green leaves on 4 Buddleja salviifolia Sage wood top and silvery white beneath. -

Cretaceous Re-Resistant Angiosperms
Cretaceous re-resistant angiosperms Shuo Wang ( [email protected] ) Qingdao University of Science and Technology https://orcid.org/0000-0003-0412-3799 Chao Shi Qingdao University of Science and Technology Hao-hong Cai Qingdao University of Science and Technology Hong-rui Zhang Qingdao University of Science and Technology Xiao-xuan Long Qingdao University of Science and Technology Erik Tihelka University of Bristol Wei-cai Song Qingdao University of Science and Technology Qi Feng Qingdao University of Science and Technology Ri-xin Jiang Qingdao University of Science and Technology Chenyang Cai Nanjing Institute of Geology and Paleontology https://orcid.org/0000-0002-9283-8323 Natasha Lombard National Herbarium, South African National Biodiversity Institute Xiong Li Kunming Institute of Botany https://orcid.org/0000-0002-1754-8721 Ji Yuan Shanghai World Expo Museum Jian-ping Zhu Shandong Normal University Hui-yu Yang Qingdao University of Science and Technology Xiao-fan Liu Qingdao University of Science and Technology Qiao-Ping Xiang Institute of Botany, Chinese Academy of Sciences Page 1/24 Zun-tian Zhao Shandong Normal University Chunlin Long Minzu University of China Xianchun Zhang The Herbarium, Institute of Botany https://orcid.org/0000-0003-3425-1011 Hua Peng Kunming Institute of Botany, CAS https://orcid.org/0000-0001-5583-537X De-Zhu Li Kunming Institute of Botany https://orcid.org/0000-0002-4990-724X Harald Schneider Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences https://orcid.org/0000- 0002-4548-7268 Michael -
Official Plant List
PINNACLE POINT OFFICIAL PLANT LIST VEGETATION PLANT LIST PINNACLE POINT. fb=fynbos th=thicket sd=sand dune dt=dune thicket mb=mossel bay area ls= limestone all=distributed in all areas MONOCOTYLEDONS Albuca sd geldbeursie; albuca; cooperi Albuca mb wittamarak; albuca; maxima Aloe arborescens th/sd kransaalwyn Aloe ferox mb tapaallwyn; opregte-aalwyn;bitteraalwyn; umhlaba(Xhosa,Zulu);ikalene,khala (Xhosa) Aleo maculata PP th bontaalwyn; spotted leafed aloe Androcymbium capense dt patrysblom; patrysie; men-in-a-boat Aristea africana fb maagbossie Aristea pusilla sd blousuurkanol Asparagus aethiopicus th katdoring;haakdoring Asparagus rubicundus th katdoring;wild asparagus; wag-n-bietjie (Asparagus asparagoides )? bridal creeper Babiana fourcadei fb bobbejaantjie Bobartia aphylla biesroei Bobartia robusta ( RARE) fb biesieblom Bolboschoenus maritimus alkali bulrush; snygras Bonatea speciosa dt moederkappie; oktoberlelie; green wood-orchid;phantom orchid Brunsvigia orientalis sd kandelaarblom Bulbine frutescens PP th rankkopieva;cat's tail;geelkatstert; balsam;Intelezi(Xhosa) Chasmanthe aethiopica dt suurkanolpypie Cymbopogon plurinodes terpentyn gras Cynodon dactylon kweek gras Disa cornuta fb inkspot disa Drimia elata dt jeukbol (eaten by Cape Grysbok) poison squill Ehrharta brevifolia naturalised weed Ehrharta bulbosa Ehrharta calycina kleinpypgras; small pipe grass Ehrharta pusilla Elegia filacea fb restio Elegia muirii (UNCOMMON) fb restio Elegia vaginulata fb dekriet Epischoenus quadrangularis N/I Eragrostis capensis ©Pinnacle Point -
The Botanic Gardens List of Rare and Threatened Species
^ JTERNATIONAL UNION FOR CONSERVATION OF NATURE AND NATURAL RESOURCES JION INTERNATIONALE POUR LA CONSERVATION DE LA NATURE ET DE SES RESSOURCES Conservation Monitoring Centre - Centre de surveillance continue de la conservation de la nature The Herbarium, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AE, U.K. BOTANIC GARDENS CONSERVATION CO-ORDINATING BODY THE BOTANIC GARDENS LIST OF RARE AND THREATENED SPECIES COMPILED BY THE THREATENED PLANTS UNIT OF THE lUCN CONSERVATION MONITORING CENTRE AT THE ROYAL BOTANIC GARDENS, KEW FROM INFORMATION RECEIVED FROM MEMBERS OF THE BOTANIC GARDENS CONSERVATION CO-ORDINATING BODY lUCN would like to express its warmest thani<s to all the specialists, technical managers and curators who have contributed information. KEW, August 198^* Tel (011-940 1171 (Threatened Plants Unit), (01)-940 4547 (Protected Areas Data Unit) Telex 296694 lUCN Secretariat: 1196 Gland, Switzerland Tel (22) 647181 Telex 22618 UNION INTERNATIONALE POUR LA CONSERVATION DE LA NATURE ET DE SES RESSOURCES INTERNATIONAL UNION FOR CONSERVATION OF NATURE AND NATURAL RESOURCES Commission du service de sauvegarde - Survival Service Commission Comite des plantes menacees — Threatened Plants Committee c/o Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AE BOTANIC GARDENS CONSERVATI6N CO-ORDINATING BODY REPORT NO. 2. THE BOTANIC GARDENS LIST OF MADAGASCAN SUCCULENTS 1980 FIRST DRAFT COMPILED BY THE lUCN THREATENED PLANTS COMMITTEE SECRETARIAT AT THE ROYAL BOTANIC GARDENS, KEW FROM INFORMATION RECEIVED FROM MEMBERS OF THE BOTANIC GARDENS CONSERVATION CO-ORDINATING BODY The TPC would like to express its warmest thanks to all the specialists, technical managers and curators who have contributed information. KEW, October, 1980 lUCN SECRETARIAT; Avenue du Mont-Blanc 1196 Gland -Suisse/Switzerland Telex: 22618 iucn Tel: (022) 64 32 54 Telegrams: lUCNATURE GLAND . -

This Article Appeared in a Journal Published by Elsevier. the Attached Copy Is Furnished to the Author for Internal Non-Commerci
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright Author's personal copy Molecular Phylogenetics and Evolution 57 (2010) 258–265 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Implications of a molecular phylogenetic study of the Malagasy genus Cedrelopsis and its relatives (Ptaeroxylaceae) Sylvain G. Razafimandimbison a,*, Marc S. Appelhans b,c, Harison Rabarison d, Thomas Haevermans e, Andriarimalala Rakotondrafara f, Stephan R. Rakotonandrasana f, Michel Ratsimbason f, Jean-Noël Labat e, Paul J.A. Keßler b,c, Erik Smets b,c,g, Corinne Cruaud h, Arnaud Couloux h, Milijaona Randrianarivelojosia i,j a Department of Botany, Bergius Foundation, Stockholm University, SE-10691, Stockholm, Sweden b Netherlands Centre for Biodiversity Naturalis (section NHN), Leiden University, 2300 RA, The Netherlands c Hortus Botanicus Leiden, Leiden, The Netherlands d Département de Biologie et Ecologie Végétales, Université d’Antananarivo, Madagascar e Muséum National d’Histoire Naturelle, Département Systématique et Evolution, UMR 7205 CNRS/MNHN Origine, Structure et Evolution de la Biodiversité, C.P.