Late Prehistoric Subsistence in the Strait of Georgia Region Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

From Semiahmoo First Nation to the Review Panel Re: Oral Presentation
Date: May 16, 2019 s EMIAHMOO FIRST NATION Canadian Environmental Assessment Agency Roberts Bank Terminal2 Project Public Hearing Phase Overview of Oral Hearing Submission by Semiahmoo First Nation To: Cindy Parker Review Panel Manager Roberts Bank Termina12 Project c/o Canadian Environmental Assessment Agency 160 Elgin Street, 22nd Floor Ottawa, ON K1A OH3 16049 Beach Road, Surrey, British Columbia, Canada V3Z 9R6 Tel: 604.536.3101 Fax: 604.536.6116 E-mail: [email protected] The Semiahmoo First Nation ("Semiahmoo") holds Aboriginal rights and title and exercises our rights, practices and culture throughout our Traditional Territory. Additionally, Semiahmoo exercises our rights, practices and culture throughout the broader resources area, which includes the lower Fraser River, Roberts Bank, Semiahmoo Bay, Boundary Bay, Fraser River, Nicomekl River, Serpentine River, Little Campbell River the Gulf Islands including San Juan Island, Vancouver Island, Washington State and the Salish Sea. Semiahmoo has communicated the adverse effects of the Roberts Band Terminal2 Project ("RBT2") on our Aboriginal rights and title to the project proponent and the Crown. Semiahmoo has previously demanded studies regarding cumulative effects of marine shipping in regard to the regarding the Marine Shipping Addendum and RBT2 including: • a traditional marine use study to examine the impacts on our Aboriginal rights and title; • a study of the effects of sedimentation on the foreshore of the Semiahmoo Indian Reserve lands from the tide, current and -

Section 12.0: Aborigin Al Consultation
ENVIRONMENTAL ASSESSMENT CERTIFICATE APPLICATION WesPac Tilbury Marine Jetty Project ABORIGINAL ABORIGINAL : 0 . 12 CONSULTATION SECTION SECTION WesPac Tilbury Marine Jetty Project Environmental Assessment Certificate Application Part C – Aboriginal Consultation Section 12.0: Aboriginal Consultation 12.0 ABORIGINAL CONSULTATION Aboriginal Interests are defined in the Section 11 Order (BCEAO, 2015b) as asserted or determined Aboriginal rights, including title, and treaty rights. An overview of planned consultation activities for the Project, activities completed to date, and a description of Aboriginal Interests is provided in Section 12.1 Aboriginal Interests. The assessment of Project-related effects on those Aboriginal Interests is presented in Section 12.1.4 Potential Effects of the Project on Aboriginal Interests. Issues raised by Aboriginal groups that do not directly relate to Aboriginal Interests, such as those pertaining to potential adverse social, economic, heritage, or health effects, and proposed measures to address those effects, are described in Section 12.2 Other Matters of Concern to Aboriginal groups. The assessment of effects on Other Matters of Concern to Aboriginal groups is also found in Section 12.2 Other Matters of Concern to Aboriginal groups. Section 12.3 provides the Issue Summary Table that summarizes Aboriginal Interests or other matters of concern to Aboriginal groups that may be affected by the Project, and the measures to avoid, mitigate or otherwise manage those effects. Information presented in this Application -

Self-Guiding Geology Tour of Stanley Park
Page 1 of 30 Self-guiding geology tour of Stanley Park Points of geological interest along the sea-wall between Ferguson Point & Prospect Point, Stanley Park, a distance of approximately 2km. (Terms in bold are defined in the glossary) David L. Cook P.Eng; FGAC. Introduction:- Geomorphologically Stanley Park is a type of hill called a cuesta (Figure 1), one of many in the Fraser Valley which would have formed islands when the sea level was higher e.g. 7000 years ago. The surfaces of the cuestas in the Fraser valley slope up to the north 10° to 15° but approximately 40 Mya (which is the convention for “million years ago” not to be confused with Ma which is the convention for “million years”) were part of a flat, eroded peneplain now raised on its north side because of uplift of the Coast Range due to plate tectonics (Eisbacher 1977) (Figure 2). Cuestas form because they have some feature which resists erosion such as a bastion of resistant rock (e.g. volcanic rock in the case of Stanley Park, Sentinel Hill, Little Mountain at Queen Elizabeth Park, Silverdale Hill and Grant Hill or a bed of conglomerate such as Burnaby Mountain). Figure 1: Stanley Park showing its cuesta form with Burnaby Mountain, also a cuesta, in the background. Page 2 of 30 Figure 2: About 40 million years ago the Coast Mountains began to rise from a flat plain (peneplain). The peneplain is now elevated, although somewhat eroded, to about 900 metres above sea level. The average annual rate of uplift over the 40 million years has therefore been approximately 0.02 mm. -

The Cordilleran Ice Sheet 3 4 Derek B
1 2 The cordilleran ice sheet 3 4 Derek B. Booth1, Kathy Goetz Troost1, John J. Clague2 and Richard B. Waitt3 5 6 1 Departments of Civil & Environmental Engineering and Earth & Space Sciences, University of Washington, 7 Box 352700, Seattle, WA 98195, USA (206)543-7923 Fax (206)685-3836. 8 2 Department of Earth Sciences, Simon Fraser University, Burnaby, British Columbia, Canada 9 3 U.S. Geological Survey, Cascade Volcano Observatory, Vancouver, WA, USA 10 11 12 Introduction techniques yield crude but consistent chronologies of local 13 and regional sequences of alternating glacial and nonglacial 14 The Cordilleran ice sheet, the smaller of two great continental deposits. These dates secure correlations of many widely 15 ice sheets that covered North America during Quaternary scattered exposures of lithologically similar deposits and 16 glacial periods, extended from the mountains of coastal south show clear differences among others. 17 and southeast Alaska, along the Coast Mountains of British Besides improvements in geochronology and paleoenvi- 18 Columbia, and into northern Washington and northwestern ronmental reconstruction (i.e. glacial geology), glaciology 19 Montana (Fig. 1). To the west its extent would have been provides quantitative tools for reconstructing and analyzing 20 limited by declining topography and the Pacific Ocean; to the any ice sheet with geologic data to constrain its physical form 21 east, it likely coalesced at times with the western margin of and history. Parts of the Cordilleran ice sheet, especially 22 the Laurentide ice sheet to form a continuous ice sheet over its southwestern margin during the last glaciation, are well 23 4,000 km wide. -

Tulalip Opening Brief
Case: 13-35773 02/28/2014 ID: 8998263 DktEntry: 16-1 Page: 1 of 110 13-35773 United States Court of Appeals for the Ninth Circuit TULALIP TRIBES Plaintiff-Appellant, v. SUQUAMISH INDIAN TRIBE Defendant-Appellee ON APPEAL FROM THE UNITED STATES DISTRICT COURT FOR THE WESTERN DISTRICT OF WASHINGTON CIVIL NO. 70-9213-RSM Subproceeding No. 05-4 TULALIP TRIBES’ OPENING BRIEF MORISSET, SCHLOSSER, JOZWIAK & SOMERVILLE Mason D. Morisset, WSBA #00273 Rebecca JCH Jackson, WSBA #44113 Email: [email protected] Email: [email protected] 801 Second Avenue, Suite 1115 Seattle, Washington 98104-1509 Tel: 206-386-5200 Fax: 206-386-7388 Attorneys for Tulalip Tribes Case: 13-35773 02/28/2014 ID: 8998263 DktEntry: 16-1 Page: 2 of 110 CORPORATE DISCLOSURE STATEMENT Appellant Tulalip Tribes is a federally recognized Indian Tribe. Accordingly, a corporate disclosure statement is not required by Federal Rule of Appellate Procedure 26.1. i Case: 13-35773 02/28/2014 ID: 8998263 DktEntry: 16-1 Page: 3 of 110 TABLE OF CONTENTS I. STATEMENT OF JURISDICTION ............................................................... 1 II. STATEMENT OF THE ISSUES .................................................................... 1 III. ADDENDUM OF PERTINENT LAWS ......................................................... 2 IV. STATEMENT OF THE CASE ....................................................................... 2 V. SUMMARY OF ARGUMENT ..................................................................... 14 VI. STANDARD OF REVIEW .......................................................................... -

Post-Glacial Sea-Level Change Along the Pacific Coast of North America Dan H
University of Washington Tacoma UW Tacoma Digital Commons SIAS Faculty Publications School of Interdisciplinary Arts and Sciences 8-1-2014 Post-glacial sea-level change along the Pacific coast of North America Dan H. Shugar University of Washington Tacoma, [email protected] Ian J. Walker Olav B. Lian Jordan BR Eamer Christina Neudorf See next page for additional authors Follow this and additional works at: https://digitalcommons.tacoma.uw.edu/ias_pub Recommended Citation Shugar, Dan H.; Walker, Ian J.; Lian, Olav B.; Eamer, Jordan BR; Neudorf, Christina; McLaren, Duncan; and Fedje, Daryl, "Post-glacial sea-level change along the Pacific oc ast of North America" (2014). SIAS Faculty Publications. 339. https://digitalcommons.tacoma.uw.edu/ias_pub/339 This Article is brought to you for free and open access by the School of Interdisciplinary Arts and Sciences at UW Tacoma Digital Commons. It has been accepted for inclusion in SIAS Faculty Publications by an authorized administrator of UW Tacoma Digital Commons. Authors Dan H. Shugar, Ian J. Walker, Olav B. Lian, Jordan BR Eamer, Christina Neudorf, Duncan McLaren, and Daryl Fedje This article is available at UW Tacoma Digital Commons: https://digitalcommons.tacoma.uw.edu/ias_pub/339 1 Post-glacial sea-level change along the Pacific coast of North 2 America 3 Dan H. Shugar1,*, Ian J. Walker1, Olav B. Lian2, Jordan B.R. Eamer1, Christina 4 Neudorf2,4, Duncan McLaren3,4, Daryl Fedje3,4 5 6 1Coastal Erosion & Dune Dynamics Laboratory, Department of Geography, 7 University of Victoria, Victoria, BC, -

Advance and Retreat of Cordilleran Ice Sheets in Washington, U.S.A
Document généré le 4 oct. 2021 19:12 Géographie physique et Quaternaire Advance and Retreat of Cordilleran Ice Sheets in Washington, U.S.A. Avancée et recul des inlandsis de la Cordillère dans l’État de Washington (É.-U.) Vorstoß und Rückzug der Kordilleren-Eisdecke in Washington State. U.S.A. Don J. Easterbrook Volume 46, numéro 1, 1992 Résumé de l'article Dans la Cordillère, les glaciations se sont produites selon des modes URI : https://id.erudit.org/iderudit/032888ar caractéristiques d'avancée et de recul : 1) dépôts fluvioglaciaires d'avancée; 2) DOI : https://doi.org/10.7202/032888ar poli glaciaire; 3) till; 4) dépôts fluvio-glaciaires de retrait au sud de Seattle, dans le sud des basses-terres de Puget, dépôts glacio-marins dans les basses-terres Aller au sommaire du numéro du nord, et eskers, terrasses fluvioglaciaires et petites moraines sur le plateau de Columbia. La datation au radiocarbone indique que les lobes de Puget et de Juan de Fuca ont avancé et reculé synchroniquement. Parmi les preuves qui Éditeur(s) nous contraignent à rejeter l'hypothèse selon laquelle un front en fusion, qui vêlait, serait à l'origine des dépôts glacio-marins, citons : 1) les nombreuses Les Presses de l'Université de Montréal datations au radiocarbone qui révèlent la mise en place simultanée de dépôts glacio-marins sur tout le territoire; 2) les dépôts issus de la fusion de la glace ISSN stagnante, intimement associés aux dépôts glacio-marins; 3) les preuves irréfutables d'une origine autre que marine des sables de Deming qui révèlent 0705-7199 (imprimé) que la Cordillère était libre de glace immédiatement avant la mise en place des 1492-143X (numérique) dépôts glacio-marins. -

Introduction
INTRODUCTION This is the fourth and final volume on the avifauna of British Columbia; it completes a discussion of the passerine birds of the province that began in Volume 3. Coming some 10 years following the publication of Volumes 1 and 2 (the non- passerine birds), it is the culmination of over 25 years of effort by the authors and significant contributions from col- leagues around the world and thousands of volunteers (Fig. 1) throughout British Columbia. The methodologies used in the preparation of this work have previously been described (Volume 1, page 146, and Volume 3, page 13) and will not be repeated here. Because of the length of time between the publication of Volumes 1 and 2 (1990) and this final volume, we have in- cluded a chapter called “Additions to the Avifauna of British Figure 2. Adult Black-legged Kittiwake with chick at nest ledge (Gjelpruvaer, Columbia, 1987 through 1999.” There we list 28 species new Norway, 11 July 1994; R. Wayne Campbell). to the province that were reported over that period. Some species, such as Manx Shearwater, Crested Caracara, Xantus’s Hummingbird, and Least Tern are new occurrence additions to the avifauna of the province. Others, such as Northern Ful- mar, Baird’s Sandpiper, Franklin’s Gull, and Black-legged Kit- tiwake (Fig. 2), now breed in the province. One species, Pa- cific Golden-Plover, has been elevated from subspecies to full species status due to taxonomic changes. Beyond the scope of the chapter are the many new changes in numbers, distribution, and results of research that contribute to our current knowledge and status of birds in the province. -

Three Dimensional Hydrostratigraphic Model of the Nanoose – Deep Bay Area, Nanaimo Lowland, British Columbia
GEOLOGICAL SURVEY OF CANADA OPEN FILE 7796 Three dimensional hydrostratigraphic model of the Nanoose – Deep Bay Area, Nanaimo Lowland, British Columbia N. Benoit, D. Paradis, J.M. Bednarski, T. Hamblin, and H.A.J. Russell 2015 GEOLOGICAL SURVEY OF CANADA OPEN FILE 7796 Three dimensional hydrostratigraphic model of the Nanoose – Deep Bay Area, Nanaimo Lowland, British Columbia N. Benoit, D. Paradis, J.M. Bednarski, T. Hamblin, and H.A.J. Russell 2015 © Her Majesty the Queen in Right of Canada, as represented by the Minister of Natural Resources Canada, 2015 doi:10.4095/296302 This publication is available for free download through GEOSCAN (http://geoscan.nrcan.gc.ca/). Recommended citation Benoit, N., Paradis, D., Bednarski, J.M., Hamblin, T., and Russell, H.A.J., 2015. Three dimensional hydrostratigraphic model of the Nanoose-Deep Bay area, Nanaimo Lowland, British Columbia; Geological Survey of Canada, Open File 7796, 1 .zip file. doi:10.4095/296302 Publications in this series have not been edited; they are released as submitted by the author. Table of Contents ABSTRACT ........................................................................................................................................................................... 1 1 INTRODUCTION ........................................................................................................................................................ 2 2 STUDY AREA ............................................................................................................................................................. -

1 a Geologic and Oceanographic Journey
Open Access e-Journal Earth Science India- www.earthscienceindia.info Popular Issue, 12 (IV), October, 2019, 1-31 A GEOLOGIC AND OCEANOGRAPHIC JOURNEY ACROSS THE SALISH SEA (SOUTHWESTERN CANADA AND NORTHWESTERN USA) Alice S. Chang and Arun Kumar The Salish Sea is also an active waterway for fisheries, industry, transportation, and recreation….. The year 2020 marks the tenth anniversary of the official naming of the Salish Sea….. The rocks of Wrangellia are mainly represented by the Karmutsen Formation, a 6,000- meter thick pile of flood basalts (flows and pillow lavas) and volcanic tuffs that were part of an oceanic plateau some 230 million years ago (late Triassic)….. Several hypotheses have been proposed to explain the origin of the Georgia Depression. These include Cretaceous–Tertiary (66 million-year-old) downwarping or down faulting of the crust, eastward tilting of the Vancouver Island block, and crustal weakening between the Insular and Coast belts. The Strait of Georgia, the Strait of Juan de Fuca, Puget Sound, and the surrounding waterways of southwestern British Columbia (BC), Canada, and northwestern Washington State, USA, are collectively called the Salish Sea (Figure 1). As one of the world’s most productive ecosystems, the Salish Sea is also an active waterway for fisheries, industry, transportation, and recreation. Scenic port cities (Vancouver and Seattle) and provincial and state capitals (Victoria and Olympia, respectively) call the shores of the Salish Sea home, as do 70% of BC’s and 60% of Washington’s populations. We decided to write this popular science article because as earth scientists, we have a long association with this region through recreation, as well as research at the universities of BC (Vancouver) and Victoria (Vancouver Island), and at Carleton University (Ottawa, Canada), where one or both of us were involved. -
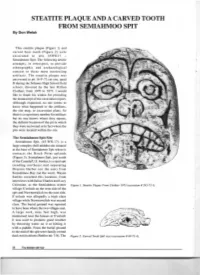
STEATITE PLAQUE and a CARVED TOOTH from SEMIAHMOO SPIT by Don Welsh
STEATITE PLAQUE AND A CARVED TOOTH FROM SEMIAHMOO SPIT By Don Welsh This steatite plaque (Figure 1) and carved bear tooth (Figure 2) were excavated in site 45WH 17 Semiahmoo Spit. The following article attempts, in retrospect, to provide ethnographic and archaeological context to these most interesting artifacts. The steatite plaque was uncovered in pit 16-F-73 cut one, quad B during the Sehome High School field school, directed by the late Milton Clothier," from 1970 to 1973. I would like to thank his widow for providing the manuscript of the excavation report. Although requested, no one seems to know what happened to the artifacts, the site map, or excavation plans. So there is a repository number for artifacts but no one knows where they repose, the definite location of the pit in which they were recovered or in fact where the pits were located within the site. The Semiahmoo Spit Site Semiahmoo Spit, ( 45-WH-17), is a large complex shell midden site situated at the base of Semiahmoo Spit where it c ontacts the Birch Point uplands (Figure 3). Semiahmoo Spit, just south of the Canada/U.S. border, is a sand spit trending northeast and separating Drayton Harbor (on the east) from Semiahmoo Bay (on the west). Wayne Suttles recorded this location, from interviews with Julius Charles and Lucy Celestine, as the Semiahmoo winter Figure 1. Steatite Plaque From Clothier 1973 (accession# 245-73-1) . village S' eel uch on the west side of the spit and Nuwnuwulich on the east side. S'eeluch was allegedly a high class village while Nuwnuwulich was second class. -

Geologic Map of the Suquamish 7.5' Quadrangle and Part of the Seattle
Geologic Map of the Suquamish 7.5’ Quadrangle and Part of the Seattle North 7.5’ x 15’ Quadrangle, Kitsap County, Washington By Ralph A. Haugerud and Kathy Goetz Troost Pamphlet to accompany Scientific Investigations Map 3181 2011 U.S. Department of the Interior U.S. Geological Survey This page intentionally left blank Contents Introduction.....................................................................................................................................................1 Geologic framework .............................................................................................................................1 About this report ...................................................................................................................................2 Stratigraphy ....................................................................................................................................................3 Glacial and nonglacial deposits .........................................................................................................3 Deposits older than Olympia interglaciation of Armstrong and others (1965) ...................3 Deposits older than Vashon stade of Fraser glaciation of Armstrong and others (1965)..............................................................................................................................4 Vashon Drift ..................................................................................................................................4 Late-glacial deposits