Soil Layer Development and Biota in Bioretention
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Basic Soil Science W
Basic Soil Science W. Lee Daniels See http://pubs.ext.vt.edu/430/430-350/430-350_pdf.pdf for more information on basic soils! [email protected]; 540-231-7175 http://www.cses.vt.edu/revegetation/ Well weathered A Horizon -- Topsoil (red, clayey) soil from the Piedmont of Virginia. This soil has formed from B Horizon - Subsoil long term weathering of granite into soil like materials. C Horizon (deeper) Native Forest Soil Leaf litter and roots (> 5 T/Ac/year are “bio- processed” to form humus, which is the dark black material seen in this topsoil layer. In the process, nutrients and energy are released to plant uptake and the higher food chain. These are the “natural soil cycles” that we attempt to manage today. Soil Profiles Soil profiles are two-dimensional slices or exposures of soils like we can view from a road cut or a soil pit. Soil profiles reveal soil horizons, which are fundamental genetic layers, weathered into underlying parent materials, in response to leaching and organic matter decomposition. Fig. 1.12 -- Soils develop horizons due to the combined process of (1) organic matter deposition and decomposition and (2) illuviation of clays, oxides and other mobile compounds downward with the wetting front. In moist environments (e.g. Virginia) free salts (Cl and SO4 ) are leached completely out of the profile, but they accumulate in desert soils. Master Horizons O A • O horizon E • A horizon • E horizon B • B horizon • C horizon C • R horizon R Master Horizons • O horizon o predominantly organic matter (litter and humus) • A horizon o organic carbon accumulation, some removal of clay • E horizon o zone of maximum removal (loss of OC, Fe, Mn, Al, clay…) • B horizon o forms below O, A, and E horizons o zone of maximum accumulation (clay, Fe, Al, CaC03, salts…) o most developed part of subsoil (structure, texture, color) o < 50% rock structure or thin bedding from water deposition Master Horizons • C horizon o little or no pedogenic alteration o unconsolidated parent material or soft bedrock o < 50% soil structure • R horizon o hard, continuous bedrock A vs. -

Soil Texture Chart Chart the Soil Texture Chart Gives Names Associated with Various Combinations of Sand, Silt and Clay and Is Used to Classify the Texture of a Soil
Name: _________________________________ Soil Texture Directions: Read, highlight, and answer the schoology questions. Until now, we have spent our time defining soil. We know that soil comes from broken down rocks and minerals that have been weathered both mechanically and chemically. We also know that soil contains a variety of parts consisting of sand, soil, clay, and humus. But what is the best soil? I’m sure you are saying loam is the best. You are right, but what exactly is loam? To understand loam, we need to understand how three parts of soil come together. If I were to ask you to draw soil, it would probably look like the image to the right. Most would just draw a pile of dirt. As we are learning though, soil is much more complex than that. Sand, Silt and Clay One of the key ways that we characterize rocks is by texture. Remember that texture refers to how something feels. It can feel grainy, rough, or smooth. Larger particles feel rough while smaller particles feel smooth. For soil, we also use texture to help characterize the type. The texture of soil, just like rocks, refers to the size of the particles that make up the soil. The terms sand, silt, and clay refer to different sizes of the soil particles. Sand, being the larger size of particles, feels gritty. Silt, being moderate in size, has a smooth or floury texture. Clay, being the smaller size of particles, feels sticky. Look at the figure to the right. The size of the circle is relative to each size particle. -

Unit 2.3, Soil Biology and Ecology
2.3 Soil Biology and Ecology Introduction 85 Lecture 1: Soil Biology and Ecology 87 Demonstration 1: Organic Matter Decomposition in Litter Bags Instructor’s Demonstration Outline 101 Step-by-Step Instructions for Students 103 Demonstration 2: Soil Respiration Instructor’s Demonstration Outline 105 Step-by-Step Instructions for Students 107 Demonstration 3: Assessing Earthworm Populations as Indicators of Soil Quality Instructor’s Demonstration Outline 111 Step-by-Step Instructions for Students 113 Demonstration 4: Soil Arthropods Instructor’s Demonstration Outline 115 Assessment Questions and Key 117 Resources 119 Appendices 1. Major Organic Components of Typical Decomposer 121 Food Sources 2. Litter Bag Data Sheet 122 3. Litter Bag Data Sheet Example 123 4. Soil Respiration Data Sheet 124 5. Earthworm Data Sheet 125 6. Arthropod Data Sheet 126 Part 2 – 84 | Unit 2.3 Soil Biology & Ecology Introduction: Soil Biology & Ecology UNIT OVERVIEW MODES OF INSTRUCTION This unit introduces students to the > LECTURE (1 LECTURE, 1.5 HOURS) biological properties and ecosystem The lecture covers the basic biology and ecosystem pro- processes of agricultural soils. cesses of soils, focusing on ways to improve soil quality for organic farming and gardening systems. The lecture reviews the constituents of soils > DEMONSTRATION 1: ORGANIC MATTER DECOMPOSITION and the physical characteristics and soil (1.5 HOURS) ecosystem processes that can be managed to In Demonstration 1, students will learn how to assess the improve soil quality. Demonstrations and capacity of different soils to decompose organic matter. exercises introduce students to techniques Discussion questions ask students to reflect on what envi- used to assess the biological properties of ronmental and management factors might have influenced soils. -

Soil Carbon Losses Due to Increased Cloudiness in a High Arctic Tundra Watershed (Western Spitsbergen)
SOIL CARBON LOSSES DUE TO INCREASED CLOUDINESS IN A HIGH ARCTIC TUNDRA WATERSHED (WESTERN SPITSBERGEN) Christoph WŸthrich 1, Ingo Mšller 2 and Dietbert Thannheiser2 1. Department of Geography, University of Basel, Spalenring 145, CH-4055 Basel, Switzerland; e-mail: [email protected]. 2. Department of Geography, University of Hamburg, Bundesstr. 55, D-20146 Hamburg, Germany. Abstract Carbon pool and carbon flux measurements of different habitats were made in the high Arctic coastal tundra of Spitsbergen. The studied catchment was situated on the exposed west coast, where westerly winds produce daily precipitation in form of rain, drizzle and fog. The storage of organic carbon in the catchment of Eidembukta amounts to 5.98 kg C m-2, mainly within the lower horizons of deep soils. Between 5.2 - 23.6 % of the carbon pool is stored in plant material. During the cold and cloudy summer of 1996, net CO2 flux measure- ments showed carbon fluxes from soil to atmosphere even during the brightest hours of the day. We estimate -2 -1 that the coastal tundra of Spitsbergen lost carbon at a rate of 0.581 g C m d predominantly as CO2-C. Carbon loss (7.625 mg C m-2 d-1) as TOC in small tundra rivers accounts only for a small proportion (1.31 %) of the total carbon loss. Introduction tetragona, Betula nana, and Empetrum hermaphroditum might accompany warming on Spitsbergen (WŸthrich, Large terrestrial carbon pools are found in the peat- 1991; Thannheiser, 1994; Elvebakk and Spjelkavik, lands of the boreal and subpolar zones that cover in 1995). -
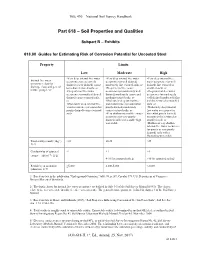
Part 618 – Soil Properties and Qualities
Title 430 – National Soil Survey Handbook Part 618 – Soil Properties and Qualities Subpart B – Exhibits 618.80 Guides for Estimating Risk of Corrosion Potential for Uncoated Steel Property Limits Low Moderate High •Very deep internal free water •Very deep internal free water •Very deep internal free Internal free water occurrence (or excessively occurrence (or well drained) water occurrence (or well occurrence class (or drained to well drained) coarse moderately fine textured soils; or drained) fine textured or drainage class) and general to medium textured soils; or •Deep internal free water stratified soils; or texture group 1/ 2/ •Deep internal free water occurrence (or moderately well •Deep internal free water occurrence (or moderately well drained) moderately coarse and occurrence (or moderately drained) coarse textured soils; medium textured soils; or well drained) moderately fine or •Moderately deep internal free and fine textured or stratified •Moderately deep internal free water occurrence (or somewhat soils; or water occurrence (or somewhat poorly drained) moderately •Moderately deep internal poorly drained) coarse textured coarse textured soils; or free water occurrence (or soils •Very shallow internal free water somewhat poorly drained) occurrence (or very poorly medium to fine textured or drained) soils with a stable high stratified soils; or water table •Shallow or very shallow internal free water occurrence (or poorly or very poorly drained) soils with a fluctuating water table Total acidity (cmol(+)/kg-1) <10 10-25 >25 3/ 4/ Conductivity of saturated <1 1-4 >4 extract (dS/m-1) 3/ 5/ 4-10 for saturated soils 6/ >10 for saturated soils 6/ Resistivity at saturation >5,000 2,000-5,000 <2,000 (ohm/cm) 1/ 7/ 1/ Based on data in the publication "Underground Corrosion," table 99, p.167, Circular 579, U.S. -

Soil Texture Key
Soil Texture Key Start Place some soil, about the size of an egg, in your palm. Add dry soil to soak up water Moisten with waterand knead until it feels like putty. yes yes Does soil remain in a no no no Is soil too dry? Is soil too wet? Sand ball when squeezed? yes Rub ball with thumb nail and note if rubbed surface shines. Place soil ball between thumb & forefinger gently pushing soil with thumb, squeezing it outward. Form a ribbon of uniform 1/8 inch thickness. Allow ribbon to emerge and extend over forefinger until it breaks under its own weight. Does the soil form a ribbon? If yes, how long? Loamy Sand no < less than 2 inch long 2 - 3 inches long; slight > 3 inches long; shines no shine when rubbed shine when rubbed when rubbed Excessively wet a pinch of soil in palm and rub with forefinger yes Sandy yes Does soil feel Sandy yes Does soil feel Sandy Does soil feel very gritty? loam very gritty? clay loam very gritty? Clay no no no Does soil feel Silty yes Does soil feel Silt yes Does soil feel Silty clay yes loam very smooth? loam very smooth? Clay very smooth? no no no Does soil feel Does soil feel Does soil feel yes Clay yes yes Loam slightly gritty Clay slightly gritty slightly gritty loam and smooth? and smooth? and smooth? Source: “Wetland Soils Genesis Hydrology, Landscape Classification” J.L. Richardson & M.J. Veprakas eds. 2001 Adapted from Thein, S.J. 1979. A flow diagram for teaching texture by-feel analysis. -

Soil Quality 1
START FARMINGSOIL QUALITY 1 introduction to soils Soil Quality Culman Steve ealthy soils yield healthy crops, But what is healthy soil and how do we Healthy, high-quality soil has: Hachieve it? • Good soil tilth • Sufficient depth Soil health is the foundation of productive farming practices. Fertile soil provides essential nutrients to • Sufficient, but not excessive, nutrient supply plants. Important physical characteristics of soil-like • Small population of plant pathogens and insect structures and aggregation allow water and air to pests infiltrate, roots to explore, and biota to thrive. Diverse • Good soil drainage and active biological communities help soil resist • Large population of beneficial organisms physical degradation and cycle nutrients at rates to • Low weed pressure meet plant needs. Soil health and soil quality are terms • No chemicals or toxins that may harm the crop used interchangeably to describe soils that are not • Resilience to degradation and unfavorable only fertile but also possess adequate physical and conditions biological properties to “sustain productivity, maintain —from Soil Health Training Manual environmental quality and promote plant and animal health” (Doron 1994). According to the (USDA) Natural Resource Remember, soil fertility is only one component Conservation Service, “Soil quality is how well soil does of soil quality. Fertile soils are able to provide the what we want it to do.” In order to grow our crops, we nutrients required for plant growth. These are the want the soil to hold water and nutrients like a sponge chemical components of soil. Some plants need certain where they are readily available for plant roots to take nutrients in large amounts, like nitrogen, phosphorus, them up, suppress pests and weeds that may attack and potassium, which are called macronutrients. -

Course Agricultural Science II Unit Soil Science Lesson Soil Texture Estimated Time Two 50-Minute Blocks Student Outcome Explain the Importance of Soil Texture
Course Agricultural Science II Unit Soil Science Lesson Soil Texture Estimated Time Two 50-minute blocks Student Outcome Explain the importance of soil texture. Learning Objectives 1. Explain the term soil texture. 2. Identify the three major separates in fine earth. 3. Explain what the different soil textures are and how they are determined. 4. Explain what rock fragments are and how they are identified. 5. Explain the term pore space. 6. Describe the importance of pore space. 7. Explain the relationship between soil texture and pore space. 8. Describe how other factors are affected by soil texture. Grade Level Expectations Resources, Supplies & Equipment, and Supplemental Information Resources 1. PowerPoint Slides PPt 1 – Flowchart for Estimating Textural Class PPt 2 – Relative Sizes of Sand, Silt, and Clay PPt 3 – Textural Triangle PPt 4 – The Soil Triangle 2. Activity Sheet AS 1 – Are All Soil Particles the Same Size? 3. Minor, Paul E. Soil Science (Student Reference). University of Missouri-Columbia: Instructional Materials Laboratory, 1995. 4. Soil Science Curriculum Enhancement. University of Missouri-Columbia: Instructional Materials Laboratory, 2003. Supplies & Equipment See AS 1 for materials and equipment needed to complete the Activity Sheet. Ag Science II – Soil Science Soil Texture • Page 1 of 7 Supplemental Information 1. Internet Sites Horizon Properties: Soil Texture. Goddard Space Flight Center, NASA. Accessed May 15, 2008, from http://soil.gsfc.nasa.gov/pvg/texture1.htm. Soil Texture. Institute of Food and Agricultural Sciences Extension, University of Florida. Accessed May 15, 2008, from http://edis.ifas.ufl.edu/SS169. Soil Texture (animation showing what kind of soils are created from different silt, sand, and clay content levels). -

5 Using the Soil Textural Triangle
Nebraska Soil Science Curriculum Using the Soil Textural Triangle Approximately 45 minutes Objectives By the end of the lesson, students will know or be able to: Use the soil textural triangle to distinguish between different types of soil Materials Preparatory Work Soil Textural Triangle Make necessary Guided Notes Page – copies 1 per student Review soil textural Guided Practice Page triangle use – 1 per student Answer key for teachers Enroll the Participants - Use the soil textural triangle to distinguish between different types of soil – Approximately 3 minute Show a large picture of a Soil Textural Triangle to the class or give each student their own copy of the triangle. Ask if anyone has seen this before or how they believe it’s used. Accept all answers until someone can identify it as a textural triangle used for determining soil texture. Provide the Experience – Approximately 5 minute Write the sample problem below on the board. Encourage students to work in small groups to determine the appropriate soil textural class. Allow groups to work together to solve the problem, wait until each group has determined the correct soil textural class. Sample Problem: 75 % Sand 15% Silt 10% Clay Answer: Sandy Loam Congratulate students for their success in using the soil textural triangle. Label the Information – Approximately 10 minute After each group has successfully identified the soil textural class explain to students that they will be writing a “How-to Guide” for this tool. In small groups, encourage students to write step by step instructions for using the soil textural triangle to accurately determine soil textural classes. -

G90-964 How Soil Holds Water
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Historical Materials from University of Nebraska-Lincoln Extension Extension 1990 G90-964 How Soil Holds Water Norman L. Klocke University of Nebraska - Lincoln Gary W. Hergert University of Nebraska - Lincoln, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/extensionhist Part of the Agriculture Commons, and the Curriculum and Instruction Commons Klocke, Norman L. and Hergert, Gary W., "G90-964 How Soil Holds Water" (1990). Historical Materials from University of Nebraska-Lincoln Extension. 725. https://digitalcommons.unl.edu/extensionhist/725 This Article is brought to you for free and open access by the Extension at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Historical Materials from University of Nebraska-Lincoln Extension by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. G90-964 How Soil Holds Water This NebGuide describes the physical characteristics that influence how soil holds water. Norman L. Klocke, Extension Water Resources Engineer Gary W. Hergert, Extension Soils Specialist z Soil Physical Characteristics z Soil Water z Available Water Capacities z Application of Soil Water Information Dryland and irrigationd agriculture depend on the management of two basic natural resources, soil and water. Soil is the supporting structure of plant life and water is essential to sustain plant life. The wise use of these resources requires a basic understanding of soil and water as well as the crop. The available water capacity and characteristics of soils are critical to water management planning for irrigationd and dryland crops. The management decisions of what crops to plant, plant populations, when to irrigation, how much to irrigation, when to apply nitrogen, and how much nitrogen to apply depend in part on the water holding capacity of soils. -

Soil Biology and Land Management
Soil Quality – Soil Biology Technical Note No. 4 Soil Biology and Land Management Table of contents The goal of soil biology management.......................................... 2 Why should land managers understand soil biology?.................. 2 The underground community....................................................... 4 What controls soil biology? ......................................................... 5 General management strategies ................................................... 7 Considerations for specific land uses......................................... 10 Cropland.............................................................................. 10 Soil Quality – Soil Biology Forestland............................................................................ 13 Technical Note No. 4 Rangeland............................................................................ 15 Assessment and monitoring....................................................... 17 January 2004 Summary.................................................................................... 19 NRCS file code: 190-22-15 References and further resources............................................... 19 Other titles in this series: #1 Soil Biology Primer Introduction #2 Introduction to Microbiotic Crusts The purpose of this technical note is to provide information about the #3 Soil Biology Slide Set effects of land management decisions on the belowground component of the food web. It points out changes in soil biological function that land managers should -

Soil Texture
Agronomy Fact Sheet Series Fact Sheet 29 Soil Texture Soil texture is an important soil characteristic in the light-textured sandy soils. The cation that drives crop production and field exchange capacity of the soil increases with management. The textural class of a soil is percent clay and organic matter (Agronomy determined by the percentage of sand, silt, Fact Sheet #22) and the pH buffering capacity and clay. Soils can be classified as one of four of a soil (its ability to resist pH change upon major textural classes: (1) sands; (2) silts; (3) lime addition), is also largely based on clay loams; and (4) clays. In this fact sheet, we will and organic matter content (Agronomy Fact discuss the importance of soil texture, different Sheet #6). Soil tilth (how easily or difficult a methods to determine soil texture, and the field is tilled) is influenced by texture, soil impact of texture on management decisions. moisture, aeration, and organic matter as well. Importance of Soil Texture Soil Textural Classes A clay soil is referred to as a fine-textured soil The combined portions of sand, silt, and clay in whereas a sandy soil is a coarse textured soil. a soil determine its textural classification. Sand Numerous soil properties are influenced by particles range in size from 0.05–2.0 mm, silt texture including: ranges from 0.002–0.05 mm, and the clay fraction is made up of particles less than 0.002 Drainage mm in diameter. Gravel or rocks greater than Water holding capacity 2 mm in diameter are not considered when Aeration determining texture.