Udayathulla April-17 Modified.Pmd
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

MCA 2Nd Year (Lateral Entry)
AweI. ky. gujrwl pMjwb tYknIkl XUnIvristI, jlMDr I. K. GUJRAL PUNJAB TECHNICAL UNIVERSITY, JALANDHAR INFORMATION BROCHURE For Academic Session : 2017-18 for Admission to 1st year (1st Semester) of “MBA & MCA” & Lateral Entry to 2nd year of MCA CORRESPONDENCE ADDRESS I.K. Gujral Punjab Technical University, Jalandhar Jalandhar – Kapurthala Highway, Kapurthala - 144603 Visit us at: www.ptu.ac.in "Technology, unlike science is a group activity. It is not based on individual intelligence, but on the interacting intelligence of many" Dr. A.P.J. Abdul Kalam Sh. Kahan Singh Pannu (IAS) : Chairman (Admissions 2017-18) Hon’ble Vice Chancellor, IKGPTU Dr. N. P. Singh : Chief Coordinator (Admissions 2017-18) -Cum- Dean (P & EP), IKGPTU Dr. Amanpreet Singh : Registrar, IKGPTU Help Desk Numbers: I. K. GUJRAL PUNJAB TECHNICAL UNIVERSITY, JALANDHAR Toll Free No. : 1800-419-0730 Mobile Nos. : +91-94780-98012, 75894-01150 E-Mail ID : [email protected] Website : www.ptu.ac.in Address : Kapurthala – Jalandhar Highway, Near Pushpa Gujral Science City, Kapurthala (Punjab), Pin Code - 144603 MAHARAJA RANJIT SINGH PUNJAB TECHNICAL UNIVERSITY, BATHINDA Contact No. : 0164-2284298 Mobile Nos. : 8725072488, 8725072339 E-Mail ID : [email protected], [email protected] Website : www.mrsptu.ac.in ATTENTION 1. The provisions in the Brochure including dates may be changed by the Competent Authority without assigning any reason. 2. For any clarification / dispute, the decision of Vice-Chancellor, I.K. Gujral Punjab Technical University Jalandhar shall be final and binding on all concerned. 3. All disputes shall be under the Jurisdiction of Jalandhar Court. CONTENTS PART – A I. K. -
Research Library Page 1
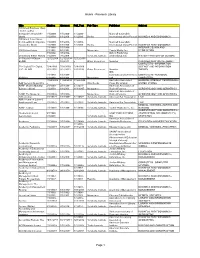
Alumni - Research Library Title Citation Abstract Full_Text Pub Type Publisher Subject 100 Great Business Ideas : from Leading Companies Around the 1/1/2009- 1/1/2009- 1/1/2009- Marshall Cavendish World 1/1/2009 1/1/2009 1/1/2009 Books International (Asia) Pte Ltd BUSINESS AND ECONOMICS 100 Great Sales Ideas : from Leading Companies 1/1/2009- 1/1/2009- 1/1/2009- Marshall Cavendish Around the World 1/1/2009 1/1/2009 1/1/2009 Books International (Asia) Pte Ltd BUSINESS AND ECONOMICS 1/1/1988- 1/1/1988- INTERIOR DESIGN AND 1001 Home Ideas 6/1/1991 6/1/1991 Magazines Family Media, Inc. DECORATION 3/1/2002- 3/1/2002- Oxford Publishing 20 Century British History 7/1/2009 7/1/2009 Scholarly Journals Limited(England) HISTORY--HISTORY OF EUROPE 33 Charts [33 Charts - 12/12/2009 12/12/2009- 12/12/2009 BLOG] + 6/3/2011 + Other Resources Newstex CHILDREN AND YOUTH--ABOUT COMPUTERS--INFORMATION 50+ Digital [50+ Digital, 7/28/2009- 7/28/2009- 7/28/2009- SCIENCE AND INFORMATION LLC - BLOG] 2/22/2010 2/22/2010 2/22/2010 Other Resources Newstex THEORY IDG 1/1/1988- 1/1/1988- Communications/Peterboro COMPUTERS--PERSONAL 80 Micro 6/1/1988 6/1/1988 Magazines ugh COMPUTERS 11/24/2004 11/24/2004 11/24/2004 Australian Associated GENERAL INTEREST PERIODICALS-- AAP General News Wire + + + Wire Feeds Press Pty Limited UNITED STATES AARP Modern Maturity; 2/1/1988- 2/1/1988- 2/1/1991- American Association of [Library edition] 1/1/2003 1/1/2003 11/1/1997 Magazines Retired Persons GERONTOLOGY AND GERIATRICS American Association of AARP The Magazine 3/1/2003+ 3/1/2003+ Magazines Retired Persons GERONTOLOGY AND GERIATRICS ABA Journal 8/1/1972+ 1/1/1988+ 1/1/1992+ Scholarly Journals American Bar Association LAW ABA Journal of Labor & Employment Law 7/1/2007+ 7/1/2007+ 7/1/2007+ Scholarly Journals American Bar Association LAW MEDICAL SCIENCES--NURSES AND ABNF Journal 1/1/1999+ 1/1/1999+ 1/1/1999+ Scholarly Journals Tucker Publications, Inc. -

Evolution of Medical Tourism in India – an Overview Himanshu Sood PCTE Group of Institutes, Ludhiana, Punjab, India
ISSN : 2348-0033 (Online) ISSN : 2249-4944 (Print) IJEAR VOL . 8, ISSU E 2, JULY - DE C 2018 Evolution of Medical Tourism in India – An Overview Himanshu Sood PCTE Group of Institutes, Ludhiana, Punjab, India Abstract high tech medical technology and western innovative methods of Medical tourism is a growing field and is of core interest of doing medication procedure at much cost effective price. So the Government of India. The industry is growing at the pace of 25% high tech equipments and upgraded medical labs are attracting and is expected to supersede from this present growth statistics. foreign nationals to take treatment from Indian premium hospitals. Every country is reporting positive sign of growth in the industry. Complicated heart surgeries, cancer care and surgeries, neuro and Even Government of India under its Ministry of Tourism has even general surgeries require high-end technology to continually formulated a separate policy for Medical Tourism. This research better outcomes, minimize complications, enable faster recovery paper is study of various aspects of Medical tourism along with and reduce length of hospital stay. initiatives taken by Government of India. It is based on secondary study and discussed various contentious issues in this Industry. C. Experienced Doctors Another advantage of taking medical treatment from India is about Keywords the experienced and well educated doctors. Many a doctors who Medical Tourism, Technology, Health and Wellness are part of industry are trained in Western countries or has taken at least some education in Western countries. This is the reason I. Introduction that they are well versed with latest equipments and understand the Travelling is an eternal concept and people move from one location use of these equipments. -

PHD September E-Bulletin 2019
PHD CHAMBER BULLETIN Vol. XLI No. 09 September-2019 ` 25 27 years of New Delhi Institute of ManagemEducatieonal Enxcellentce Estd 1992 | Approved by AICTE since 1996 | Declared equivalent to MBA by AICTE in 2008 | Among “Illustrious Brands Making In dia Proud 2017”- Mail Today | Among “The Fastest Growing Business Schools of Asia”- process reviewer - KPMG - India | Among “The World’s Greatest Brands 2015-16 & 2016-17 In dia in Education Category”- process reviewer - PwC | In ternationally Accredited by ASIC, U.K. Since 2014 as a Premier B-School BEST MBA SCHOOL IN INDIA FOR PLACEMENT SERVICES : ASSOCHAM 2018 | MOST PREFERRED B-SCHOOL OF THE YEAR-NORTH : ASSOCHAM 2017 AND 2019 RATED 2017 & 2018 RANKINGS OF AICTE (GOVT. OF INDIA) & CII BE ST ‘Best Industry Linked Management School of India’-2017 B Y IN ‘Best Industry Linked Management School of India’-2018 D US T R Y & N A T ION A L T R ADE C H A MB ER S CI I A ND A SSOC H A M | NDIM thanks Mr. Arun Jaitley, Bharat Ratna Hon'ble Pranab Mukherjee, Hon'ble Dr. Manmohan Singh, Mr. Prakash Javadekar, PA Mr. Harshvardhan Neotia, Col. Rajyavardhan Singh Rathore, Mr. Suresh Prabhu, Mr. Sunil Bharti Mittal, Mr. C.P. Gurnani, Mr. Suresh R Narayanan, Mr. Analjit Singh, Mr. Subhash Chandra, Dr. Niranjan Hiranandani, Mr. Anil Khaitan, Mr. Kanwaljeet Jawa, Dr. Mahesh Gupta, TN Mr. Hemant Kanoria, Mr. Deepak Kapoor, Dr. Arvind Lal, Mr. Sunil Duggal, Mr. Anurag Batra, Mr. Shekhar Gupta, Mr. Rajat Sharma, ER Mr. Anurag Thakur, Mr. Suresh Kumar, Mr. -

Vol 3, Issue 1
A Bi-annual International Journal ISSN 2349 - 1078 Jan - July 2014 Vol: 3 Issue 1 ADVISORY BOARD: CONTENTS • Dr. B.K Mohanty, Professor, IIM Lucknow. 1 - Factors Influencing Online Shopping Behaviour: A Study of Indian and Australian 4 - 14 Customers • Prof. Deepak Sharma, Associate Dean, International Director Centre for Energy Policy, Australia. Dr. GunmalaSuri, Assistant Professor, UBS, Panjab University, Chandigarh. • Mr. Ativ Ralli, Vice-President, Deutsche Bank. Dr. RupinderBir Kaur, Assistant Professor, UBS, Panjab University, Chandigarh. • Mr. Bobinder Makkar, Director, Product Management, CISCO Head Office, America. 2 - Systematic approach to Value Engineering and Cost Reduction: A case study on Indian 15 - 28 • Mr. Sushil Sachdev, Vice-President, Eastman Industries Ltd., Ludhiana. Automobile Industries Dr. K. K. Patra, Professor of Finance and Dean, Rourkela Institute of Management Studies, • Dr. Baltej Singh Mann, Professor, Sanjose State University, USA. Rourkela. • Dr. Sanjit Dhami, Professor, Department of Economics, University of Leicester, UK. 3 - A Critical Review of Initiatives Taken by IT and Electronics Companies in India to 29 - 37 • Dr. Sanjeev Sofat, Professor and Head, PEC University of Technology, Chandigarh. Control E-Waste • Mr. Dirk Leysen, Professor and Economist, KHK Kemphen University College, Belgium. Dr. Abhiruchi Singh Verma, Associate Prof., Institute of I.T &Management, New Delhi. 4 - Status Of Rural Female Work Force Participation Rates In India 38 - 44 • Dr. Dr. G.S. Batra, Professor, School of Management Studies, Punjabi University, Pataila. Dr. D. Pulla Rao, Professor of Economics, Andhra University, Visakhapatnam, A.P. • Dr. D.D. Arora, Dean, Colleges, Kurukshetra University, Kurukshetra. 5 - New Dimensions in Service Sector – Banking, Insurance and Transport 49 - 58 • Mr. -

Vol. 7 | June 2020
Vol. 7 | June 2020 Biannual Magazine - Centre for BRICS Studies RUSSIA CHINA INDIA BRAZIL SOUTH AFRICA BUSINESS IN BRICS VOL: 7 I JUNE 2020 VISION OF HONOURABLE CHANCELLOR The Centre for BRICS Studies was inaugurated on February 10, 2011 at Amity University Haryana by Dr. Aseem Chauhan, Honourable Chancellor. His dynamic leadership has constantly been a source of inspiration in strengthening and transforming the “Centre for BRICS Studies” into a premier Centre of Excellence in Amity University Haryana promoting high-end research, practice and outreach. The Chancellor’s rightful vision of BRICS becoming an important geo-political and geo- economic grouping in the arena of multilateral diplomacy, business and socio-cultural relations entailing extensive research and academic exploration for a large university like Amity University Haryana has been truly a catalytic force in making the “Centre for BRICS Studies” a hub for research, knowledge creation and dissemination. The launching of 7th Volume of the Centre’s Newsletter in the wake of global pandemic reiterates the commitment of the Centre’s endeavour to promote the spirit of academic research through academia-industry-society linkage in conjunction with relentless effort towards innovation and capacity building while fighting the global pandemic crisis successfully, which in turn resonates the vision and charismatic leadership of Hon’ble Chancellor Dr. Aseem Chauhan. Dr. Aseem Chauhan Chancellor, Amity University Haryana 2 BUSINESS IN BRICS VOL: 7 I JUNE 2020 ABOUT AMITY UNIVERSITY HARYANA Amity University Gurugram is a premier research and innovation driven university in India providing its student with world class academic and learning environment. Spread over a sprawling campus of 110 acres and located in the millennium city of Gurgaon, Amity University Gurgaon has become a stellar destination for students from across the country and abroad. -
PCTE Brochure
D. 1999 Business Management I Commerce I Computer Applications I Hotel Management I Pharmacy I Travel & Tburism I Biotechnology I Medical Lab Sciences I Agriculture (Honors) I Fashion Design I Journalism & Mass Communication I ?a*t7tuO;r,ornuGu*uo/ PCTE group of tnstitutes is one of the most dynamic, Vibrant and encompassing amongst all the colleges in North lndia. PCTE Group of lnstitutes recognizes that today's world requires young men and women who not only have knowledge about the modern technologies and business techniques but also have practical knowledge to use these technologies. PCTE has consciously made efforts to give latest and practical knowledge to its students. PCTE is an approved institution by All lndia Council of Technical Education (AICTE), Ministry of HRD Govt. of lndia and affiliated to Punjab Technical University, Jalandhar. "You con apply altthat knowledge backto the business and -44' have a securefuture". Dr. K.N.S. Kang Chairman cum Director General B.E. (Elect), MBA, Ph.D (Mktg.) ABSTRACT PCTE has students & Alumni from 50 countries QuickFacts including lndia. Founded in 1999 a 1Oth best B-School in the country on ROI by Outlook . 2glnternational&NationalAwards 2017 including llM's. & . 43 UniversityGold Medals a Active Students exchange programs with France Lithuania . Ranked Best B-School in Punjab, HB J&K and Rajasthan 33 Ambassadors & Seniors Delegates of various in last the decade by various national magazines and cou ntries have visited PCTE. Journals, including Outlook, Business lndia, AIMA, etc' a 125 companies recruited students of MBfuMBA-lB students . 24Olnternational from campus for Batch 201 6 -1 8. -

October 2019 Management Times
AIMA’S MONTHLY E-MAGAZINE AIMA | SNAPSHOTS | OCTOBER 2019 MANAGEMENT TIMES OCTOBER 2019 AIMA OFFICE BEARERS PRESIDENT Mr Sanjay Kirloskar Chairman & Managing Director Dear Readers, Kirloskar Brothers Ltd It gives me great pleasure to present the October issue of AIMA News. With your support and encouragement AIMA continues to strive to spread management SENIOR VICE PRESIDENT thought and build capability through its various initiatives and programmes during the period. Mr Harsh Pati Singhania AIMA participated in the Asian Association of Management Organisation’s (AAMO) Director, JK Organisation and 2nd Council Meeting of 2019 and 8th Annual General Meeting held on 23-25 Vice Chairman & Managing Director October 2019 in Sydney, Australia. AAMO representatives, including those from JK Paper Ltd Macau, India, Australia, Malaysia, Philippines, Nepal and others, gathered to discuss the current and future plans of the organisation. AAMO is a partnership of NMOs in VICE PRESIDENT the Asian Pacific region whose purpose is to share and actively leverage resources to enhance the achievement of their respective missions. The current 11 members Mr C K Ranganathan of AAMO include Australia, Hong Kong, India, Macau, Malaysia, Nepal, New Zealand, Chairman & Managing Director Pakistan, Philippines, Sri Lanka and Korea. CavinKare Pvt Ltd India (AIMA) took over the Presidency of AAMO from Malaysia at this meeting and nominated Mr Rajive Kaul, Past President, AIMA & Chairman, NICCO Group as TREASURER President. The AAMO Secretariat which rotates with the Presidency will now be Mr Nikhil Sawhney handled by AIMA for the next three years. AIMA is an important and long-time member of AAMO, and looks forward to your continued support as it dons the Vice Chairman and Managing Director mantle of AAMO Presidency for the fourth time. -

Chinese, Indian Troops Begin Pullback Process
brightpunjabexpress.com CHANDIGARH > FRIDAY, FEBRUARY 12, 2021 RNI.No. PUNENG/2018/76070 | VOLUME 3, ISSUE 162 | `3.00 CAPT ENJOYS CONFIDENCE OF EVERY HEALTH DEPT ISSUES 12 CHALLANS TO PITCH FOR 2ND TEST EXPECTED PUNJABI: BRAHM MOHINDRA P03 VENDORS SELLING TOBACCO PRODUCTS P05 TO TURN FROM DAY 1 P12 Chinese, Indian troops BJP doesn’t believe in political untouchability: PM NEW DELHI, FEBRUARY 11 begin pullback process Prime Minister Narendra Modi on Thursday asserted that the BJP does not be- lieve in political untouch- CHINA TO MOVE TROOPS TO FINGER 8 FROM FINGER 4: RAJNATH “It was also agreed to hold the next meeting of Senior ability and values consen- NEW DELHI, FEBRUARY 11 Commanders of the two armies sus in politics, as he noted within 48 hours after complete that his government has India and China have disengagement in Pangong lake bestowed state honours on reached an agreement on areas to resolve any remaining even the ruling party’s po- disengagement in the North issues. In addition, the Chinese litical rivals to acknowledge and South banks of Pan- side had mobilised a large their service to the nation. Prime Minister Narendra Modi speaks during ‘Samarpan gong lake in eastern Ladakh number of troops and arma- Addressing Bharatiya Diwas’ event, organised to pay homage to former Bharatiya that mandates both sides ments along the LAC as well as Janata Party (BJP) MPs Jana Sangh leader Pandit Deendayal Upadhyaya on his to “cease” forward deploy- in the depth areas. Our armed at an event here to mark 53rd death anniversary, in New Delhi, on Thursday. -

Ik Gujral Punjab Technical University Part-A
PART-A I. K. GUJRAL PUNJAB TECHNICAL UNIVERSITY ADMINISTRATIVE BLOCK OF I. K. GUJRAL PUNJAB TECHNICAL UNIVERSITY, JALANDHAR 1. ABOUT I. K. GUJRAL PUNJAB TECHNICAL UNIVERSITY Punjab Technical University (PTU) was established in the year 1997 under The Punjab Technical University Act, 1996 (Punjab Act No. 1 of 1997) An Act to provide for the establishment and incorporation of a University for the advancement of technical education and development thereof in the State of Punjab and for matters connected therewith. PTU was renamed as I. K. Gujral Punjab Technical University (IKGPTU) through THE PUNJAB TECHNICAL UNIVERSITY (AMNDMENT) BILL, 2014 vide Bill No. 17-PLA-2014 dated 17th Jul, 2014. As per amended Act the jurisdiction of following Districts of Punjab:- 1. Amritsar 5. Kapurthala 9. Rupnagar 2. Gurdaspur 6. Ludhiana 10. S.A.S. Nagar, Mohali 3. Hoshiarpur 7. Nawanshahr 4. Jalandhar 8. Pathankot After winning the Best Technical University Award 2013-14 from ISTE, India, now this year, IKGPTU has been recognized as the 3rd best university in the state by MHRD, India. Furthermore, in this Framework 2016, the university got 33rd position for its best research contributions and 54th position in top 100 best universities of the country. It is indeed a matter of pride for all of us. In pursuance of Punjab Act No. 1 of 1997, the University started its journey with 09 engineering colleges and few management colleges affiliated to it. Since then the University has come a long way and at present the University, under its administration, has approx 186 colleges offering courses in Engineering, Management, Pharmacy, B.Sc.