Speciation Without Changes in Genital Shape: a Case Study on Brazilian Pholcid Spiders (Araneae: Pholcidae) Bernhard A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Aranhas (Araneae, Arachnida) Do Estado De São Paulo, Brasil: Diversidade, Esforço Amostral E Estado Do Conhecimento
Biota Neotrop., vol. 11(Supl.1) Aranhas (Araneae, Arachnida) do Estado de São Paulo, Brasil: diversidade, esforço amostral e estado do conhecimento Antonio Domingos Brescovit1,4, Ubirajara de Oliveira2,3 & Adalberto José dos Santos2 1Laboratório de Artrópodes, Instituto Butantan, Av. Vital Brasil, n. 1500, CEP 05503-900, São Paulo, SP, Brasil, e-mail: [email protected] 2Departamento de Zoologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais – UFMG, Av. Antonio Carlos, n. 6627, CEP 31270-901, Belo Horizonte, MG, Brasil, e-mail: [email protected], [email protected] 3Pós-graduação em Ecologia, Conservação e Manejo da Vida Silvestre, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais – UFMG 4Autor para correspondência: Antonio Domingos Brescovit, e-mail: [email protected] BRESCOVIT, A.D., OLIVEIRA, U. & SANTOS, A.J. Spiders (Araneae, Arachnida) from São Paulo State, Brazil: diversity, sampling efforts, and state-of-art. Biota Neotrop. 11(1a): http://www.biotaneotropica.org. br/v11n1a/en/abstract?inventory+bn0381101a2011. Abstract: In this study we present a database of spiders described and registered from the Neotropical region between 1757 and 2008. Results are focused on the diversity of the group in the State of São Paulo, compared to other Brazilian states. Data was compiled from over 25,000 records, published in scientific papers dealing with Neotropical fauna. These records enabled the evaluation of the current distribution of the species, the definition of collection gaps and priority biomes, and even future areas of endemism for Brazil. A total of 875 species, distributed in 50 families, have been described from the State of São Paulo. -

Arachnida, Solifugae) with Special Focus on Functional Analyses and Phylogenetic Interpretations
HISTOLOGY AND ULTRASTRUCTURE OF SOLIFUGES Comparative studies of organ systems of solifuges (Arachnida, Solifugae) with special focus on functional analyses and phylogenetic interpretations HISTOLOGIE UND ULTRASTRUKTUR DER SOLIFUGEN Vergleichende Studien an Organsystemen der Solifugen (Arachnida, Solifugae) mit Schwerpunkt auf funktionellen Analysen und phylogenetischen Interpretationen I N A U G U R A L D I S S E R T A T I O N zur Erlangung des akademischen Grades doctor rerum naturalium (Dr. rer. nat.) an der Mathematisch-Naturwissenschaftlichen Fakultät der Ernst-Moritz-Arndt-Universität Greifswald vorgelegt von Anja Elisabeth Klann geboren am 28.November 1976 in Bremen Greifswald, den 04.06.2009 Dekan ........................................................................................................Prof. Dr. Klaus Fesser Prof. Dr. Dr. h.c. Gerd Alberti Erster Gutachter .......................................................................................... Zweiter Gutachter ........................................................................................Prof. Dr. Romano Dallai Tag der Promotion ........................................................................................15.09.2009 Content Summary ..........................................................................................1 Zusammenfassung ..........................................................................5 Acknowledgments ..........................................................................9 1. Introduction ............................................................................ -

Zootaxa, a Review of the Cellar Spider Genus Psilochorus Simon 1893 In
Zootaxa 2144: 1–53 (2009) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2009 · Magnolia Press ISSN 1175-5334 (online edition) A review of the cellar spider genus Psilochorus Simon 1893 in America north of Mexico (Araneae: Pholcidae) JOZEF SLOWIK University of Alaska Museum of the North, Department of Entomology, 907 Yukon Drive, Fairbanks, Alaska 99775 USA. E-mail: [email protected] Table of contents Abstract ............................................................................................................................................................................... 2 Introduction ......................................................................................................................................................................... 2 Materials and methods ........................................................................................................................................................ 2 Taxonomy ............................................................................................................................................................................ 3 Pholcidae C. L. Koch, 1851 ................................................................................................................................................ 3 Psilochorus Simon, 1893 .................................................................................................................................................... 3 Key to Psilochorus species in America -
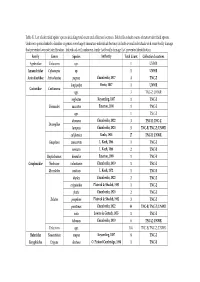
Table S1. List of Identified Spider Species Including Total Count and Collection Locations. Bold Cells Include Counts of Mature Identified Species
Table S1. List of identified spider species including total count and collection locations. Bold cells include counts of mature identified species. Unknown species linked to families or genera were largely immature individuals but may include several individuals with some bodily damage that prevented accurate identification. Individuals with unknown family had bodily damage that prevented identification. Family Genus Species Authority Total Count Collection Locations Agelenidae Unknown spp. 1 UNWR Amaurobiidae Cybaeopsis sp. 1 UNWR Antrodiaetidae Antrodiaetus pugnax Chamberlin, 1917 4 TNC-Z longipalpa Hentz, 1847 1 UNWR Corinnidae Castianeira spp. 3 TNC-Z; UNWR neglectus Keyserling, 1887 1 TNC-Z Drassodes saccatus Emerton, 1890 1 TNC-Z spp. 1 TNC-Z dromeus Chamberlin, 1922 3 TNC-B; TNC-Z Drassyllus lamprus Chamberlin, 1920 3 TNC-B; TNC-Z; UNWR californica Banks, 1904 17 TNC-B; UNWR Gnaphosa muscorum L. Koch, 1866 1 TNC-Z sericata L. Koch, 1866 2 TNC-B Haplodrassus hiemalis Emerton, 1909 1 TNC-B Gnaphosidae Nodocion voluntaries Chamberlin, 1919 1 TNC-Z Urozelotes rusticus L. Koch, 1872 1 TNC-B duplex Chamberlin, 1922 2 TNC-Z exiguioides Platnick & Shadab, 1983 1 TNC-Z fratis Chamberlin, 1920 2 TNC-Z Zelotes josephine Platnick & Shadab, 1983 3 TNC-Z puritanus Chamberlin, 1922 44 TNC-B; TNC-Z; UNWR sula Lowrie & Gertsch, 1955 1 TNC-Z tubuous Chamberlin, 1919 6 TNC-Z; UNWR Unknown spp. 184 TNC-B; TNC-Z; UNWR Hahniidae Neoantistea magna Keyserling, 1887 8 TNC-Z Linyphiidae Erigone dentosa O. Pickard-Cambridge, 1894 1 TNC-B spp. 1 TNC-Z Unknown spp. 13 TNC-Z; UNWR mccooki Montgomery, 1904 95 TNC-B; TNC-Z; UNWR Schizocosa minnesotensis Gertsch, 1934 25 TNC-B Lycosidae spp. -

SA Spider Checklist
REVIEW ZOOS' PRINT JOURNAL 22(2): 2551-2597 CHECKLIST OF SPIDERS (ARACHNIDA: ARANEAE) OF SOUTH ASIA INCLUDING THE 2006 UPDATE OF INDIAN SPIDER CHECKLIST Manju Siliwal 1 and Sanjay Molur 2,3 1,2 Wildlife Information & Liaison Development (WILD) Society, 3 Zoo Outreach Organisation (ZOO) 29-1, Bharathi Colony, Peelamedu, Coimbatore, Tamil Nadu 641004, India Email: 1 [email protected]; 3 [email protected] ABSTRACT Thesaurus, (Vol. 1) in 1734 (Smith, 2001). Most of the spiders After one year since publication of the Indian Checklist, this is described during the British period from South Asia were by an attempt to provide a comprehensive checklist of spiders of foreigners based on the specimens deposited in different South Asia with eight countries - Afghanistan, Bangladesh, Bhutan, India, Maldives, Nepal, Pakistan and Sri Lanka. The European Museums. Indian checklist is also updated for 2006. The South Asian While the Indian checklist (Siliwal et al., 2005) is more spider list is also compiled following The World Spider Catalog accurate, the South Asian spider checklist is not critically by Platnick and other peer-reviewed publications since the last scrutinized due to lack of complete literature, but it gives an update. In total, 2299 species of spiders in 67 families have overview of species found in various South Asian countries, been reported from South Asia. There are 39 species included in this regions checklist that are not listed in the World Catalog gives the endemism of species and forms a basis for careful of Spiders. Taxonomic verification is recommended for 51 species. and participatory work by arachnologists in the region. -

Volume 28, No. 2, Fall 2009
Fall 2009 Vol. 28, No. 2 NEWSLETTER OF THE BIOLOGICAL SURVEY OF CANADA (TERRESTRIAL ARTHROPODS) Table of Contents General Information and Editorial Notes ..................................... (inside front cover) News and Notes News from the Biological Survey of Canada ..........................................................27 Report on the first AGM of the BSC .......................................................................27 Robert E. Roughley (1950-2009) ...........................................................................30 BSC Symposium at the 2009 JAM .........................................................................32 Demise of the NRC Research Press Monograph Series .......................................34 The Evolution of the BSC Newsletter .....................................................................34 The Alan and Anne Morgan Collection moves to Guelph ......................................34 Curation Blitz at Wallis Museum ............................................................................35 International Year of Biological Diversity 2010 ......................................................36 Project Update: Terrestrial Arthropods of Newfoundland and Labrador ..............37 Border Conflicts: How Leafhoppers Can Help Resolve Ecoregional Viewpoints 41 Project Update: Canadian Journal of Arthropod Identification .............................55 Arctic Corner The Birth of the University of Alaska Museum Insect Collection ............................57 Bylot Island and the Northern Biodiversity -

Pholcid Spiders from the Lower Guinean Region of Central Africa: an Overview, with Descriptions of Seven New Species (Araneae, Pholcidae)
European Journal of Taxonomy 81: 1-46 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.81 www.europeanjournaloftaxonomy.eu 2014 · Huber B.A. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Research article urn:lsid:zoobank.org:pub:AC69F89F-C11B-49B1-8EEE-183286EDA755 Pholcid spiders from the Lower Guinean region of Central Africa: an overview, with descriptions of seven new species (Araneae, Pholcidae) Bernhard A. HUBER1,5, Philippe LE GALL2,6, Jacques François MAVOUNGOU3,4,7 1 Alexander Koenig Research Museum of Zoology, Adenauerallee 160, 53113 Bonn, Germany E-mail: [email protected] 2 Laboratoire Evolution, Génomes et Spéciation, UPR 9034, Centre National de la Recherche Scientifique (CNRS), 91198 Gif sur Yvette Cedex, France and Université Paris-Sud 11, 91405 Orsay Cedex, France. E-mail: [email protected] 3 Institut de Recherche en Ecologie Tropicale, BP: 13354, Libreville, Gabon Email: [email protected] 4 Université des Sciences et Techniques de Masuku, Franceville, Gabon. 5 urn:lsid:zoobank.org:author:33607F65-19BF-4DC9-94FD-4BB88CED455F 6 urn:lsid:zoobank.org:author:13F0CC41-6013-49FD-B4C2-0A455C9F8D82 7 urn:lsid:zoobank.org:author:E990D817-154C-4B8A-BD6D-740B05879DA0 Abstract. This paper summarizes current knowledge about Central African pholcids. Central Africa is here defined as the area between 10°N and 7°S and between 6°E and 18°E, including mainly the Lower Guinean subregion of the Guineo-Congolian center of endemism. This includes all of Gabon, Equatorial Guinea, São Tomé and Príncipe, most of Cameroon and Congo Republic, and parts of the neighboring countries. -

Bonn Zoological Bulletin Volume 58 Pp
0 © Biodiversity Heritage Library, http://www.biodiversitylibrary.org/; www.zoologicalbulletin.de; www.biologiezentrum.at Bonn zoological Bulletin Volume 58 pp. 217-226 Bonn, November 20 1 Non-insect arthropod types in the ZFMK collection, Bonn (Acari, Araneae, Scorpiones, Pantopoda, Am phi pod a) Bernhard A. Huber & Stefanie Lankhorst Zoologisches Forschungsmuseum Alexander Koenig, Adenauerallee 1 60, D-53 1 1 3 Bonn, Germany; E-mail: [email protected] Abstract. The type specimens of Acari, Araneae, Scorpiones, Pantopoda, and Amphipoda housed in the Alexander Koenig Zoological Research Museum, Bonn, are listed. 183 names are recorded; of these, 64 (35%) are represented by name bearing (i.e., primary) types. Specific and subspecific names are listed alphabetically, followed by the original genus name, bibliographic citation, present combination (as far as known to the authors), and emended label data. Key Words. Type specimens, Acari, Araneae, Scorpiones, Pantopoda, Amphipoda, Bonn. INTRODUCTION The ZFMK in Bonn has a relatively small collection of Abbreviations. HT: holotype, PT: paratype, ST: syntype, non-insect arthropods, with an emphasis on arachnids LT: lectotype, PLT: paralectotype; n, pn, dn, tn: (proto-, (mostly mites, spiders, and scorpions), sea spiders (Pan- deuto-, trito-) nymph, hy: hypopus, L: larva topoda) and amphipods. Other arachnid and crustacean or- ders are represented, but not by type material. A small part of the material goes back to the founder of the museum, ACARI Alexander Koenig, and was collected around 1910. Most Acari were deposited at the museum by F. S. Lukoschus aequatorialis [Orycteroxemis] Lukoschus, Gerrits & (mostly Astigmata: Glyciphagidae, Atopomelidae, etc.), Fain, 1977b. PT,' 2 slides. -

Bromeliads As Biodiversity Amplifiers and Habitat Segregation of Spider Communities in a Neotropical Rainforest
2010. The Journal of Arachnology 38:270–279 Bromeliads as biodiversity amplifiers and habitat segregation of spider communities in a Neotropical rainforest Thiago Gonc¸alves-Souza1, Antonio D. Brescovit2, Denise de C. Rossa-Feres1,andGustavo Q. Romero1,3: 1Departamento de Zoologia e Botaˆnica, IBILCE, Universidade Estadual Paulista, UNESP, Rua Cristo´va˜o Colombo 2265, CEP 15054- 000, Sa˜o Jose´ do Rio Preto, SP, Brazil; 2Instituto Butanta˜, Laborato´rio de Artro´podes Pec¸onhentos, Avenida Vital Brazil 1500, CEP 05503-900, Sa˜o Paulo, SP, Brazil Abstract. Although bromeliads can be important in the organization of invertebrate communities in Neotropical forests, few studies support this assumption. Bromeliads possess a three-dimensional architecture and rosette grouped leaves that provide associated animals with a good place for foraging, reproduction and egg laying, as well as shelter against desiccation and natural enemies. We collected spiders from an area of the Atlantic Rainforest, southeastern Brazil, through manual inspection in bromeliads, beating trays in herbaceous+shrubby vegetation and pitfall traps in the soil, to test if: 1) species subsets that make up the Neotropical forest spider community are compartmentalized into different habitat types (i.e., bromeliads, vegetation and ground), and 2) bromeliads are important elements that structure spider communities because they generate different patterns of abundance distributions and species composition, and thus amplify spider beta diversity. Subsets of spider species were compartmentalized into three habitat types. The presence of bromeliads represented 41% of the increase in total spider richness, and contributed most to explaining the high beta diversity values among habitats. Patterns of abundance distribution of the spider community differed among habitats. -

Epigeic Spider (Araneae) Diversity and Habitat Distributions in Kings
Clemson University TigerPrints All Theses Theses 5-2011 Epigeic Spider (Araneae) Diversity and Habitat Distributions in Kings Mountain National Military Park, South Carolina Sarah Stellwagen Clemson University, [email protected] Follow this and additional works at: https://tigerprints.clemson.edu/all_theses Part of the Entomology Commons Recommended Citation Stellwagen, Sarah, "Epigeic Spider (Araneae) Diversity and Habitat Distributions in Kings Mountain National Military Park, South Carolina" (2011). All Theses. 1091. https://tigerprints.clemson.edu/all_theses/1091 This Thesis is brought to you for free and open access by the Theses at TigerPrints. It has been accepted for inclusion in All Theses by an authorized administrator of TigerPrints. For more information, please contact [email protected]. EPIGEIC SPIDER (ARANEAE) DIVERSITY AND HABITAT DISTRIBUTIONS IN KINGS MOUNTAIN NATIONAL MILITARY PARK, SOUTH CAROLINA ______________________________ A Thesis Presented to the Graduate School of Clemson University _______________________________ In Partial Fulfillment of the Requirements for the Degree Masters of Science Entomology _______________________________ by Sarah D. Stellwagen May 2011 _______________________________ Accepted by: Dr. Joseph D. Culin, Committee Chair Dr. Eric Benson Dr. William Bridges ABSTRACT This study examined the epigeic spider fauna in Kings Mountain National Military Park. The aim of this study is to make this information available to park management for use in the preservation of natural resources. Pitfall trapping was conducted monthly for one year in three distinct habitats: riparian, forest, and ridge-top. The study was conducted from August 2009 to July 2010. One hundred twenty samples were collected in each site. Overall, 289 adult spiders comprising 66 species were collected in the riparian habitat, 345 adult comprising 57 species were found in the forest habitat, and 240 adults comprising 47 species were found in the ridge-top habitat. -

Ana Letícia Trivia Diversidade De Aranhas (Arachnida, Araneae) De
Ana Letícia Trivia Diversidade de aranhas (Arachnida, Araneae) de solo na Mata Atlântica do Parque Municipal da Lagoa do Peri, Florianópolis, SC, Brasil Trabalho submetido à disciplina BIO7016 - Trabalho de Conclusão de Curso II da Universidade Federal de Santa Catarina, requisito parcial para a obtenção do Grau de Bacharel em Ciências Biológicas. Orientador: Dr. Benedito Cortês Lopes Co-orientador: Dr. Amazonas Chagas Júnior. Florianópolis (SC) Fevereiro de 2013 i Ana Letícia Trivia Diversidade de aranhas (Arachnida, Araneae) de solo na Mata Atlântica do Parque Municipal da Lagoa do Peri, Florianópolis, SC, Brasil Este Trabalho de Conclusão de Curso foi julgado e adequado para obtenção de Título de Bacharel em Ciências Biológicas, aprovado em sua forma final pelo Curso de Ciências Biológicas da Universidade Federal de Santa Catarina Florianópolis (SC), 20 de Fevereiro de 2013. _________________________________________ Professora Maria Risoleta Freire Marques Coordenadora do Curso _________________________________________ Professora Kieiv Resende Souza de Moura Subcoordenadora Banca Examinadora: _________________________________________ Professor Dr. Benedito Cortês Lopes Universidade Federal de Santa Catarina _________________________________________ Professor Dr. Amazonas Chagas Júnior Universidade Federal do Mato Grosso _________________________________________ Professora Dra. Malva Isabel Medina Hernández Universidade Federal de Santa Catarina _________________________________________ Professor Dr. Luiz Carlos de Pinho Universidade Federal de Santa Catarina ii Ao meu grande irmão, por tudo que me ensinou, por tudo que fez por mim e pelo imenso apoio em todas as dificuldades. iii AGRADECIMENTOS Meus sinceros agradecimentos a todos que me apoiaram no decorrer deste trabalho, direta ou indiretamente, ajudando não somente no desenvolvimento do trabalho, mas como pessoa, estando ao meu lado ao longo da minha vida e da minha graduação. -

Pinyon Pine Mortality Alters Communities of Ground-Dwelling Arthropods
Western North American Naturalist 74(2), © 2014, pp. 162–184 PINYON PINE MORTALITY ALTERS COMMUNITIES OF GROUND-DWELLING ARTHROPODS Robert J. Delph1,2,6, Michael J. Clifford2,3, Neil S. Cobb2, Paulette L. Ford4, and Sandra L. Brantley5 ABSTRACT.—We documented the effect of drought-induced mortality of pinyon pine (Pinus edulis Engelm.) on com- munities of ground-dwelling arthropods. Tree mortality alters microhabitats utilized by ground-dwelling arthropods by increasing solar radiation, dead woody debris, and understory vegetation. Our major objectives were to determine (1) whether there were changes in community composition, species richness, and abundance of ground-dwelling arthro- pods associated with pinyon mortality and (2) whether specific habitat characteristics and microhabitats accounted for these changes. We predicted shifts in community composition and increases in arthropod diversity and abundance due to the presumed increased complexity of microhabitats from both standing dead and fallen dead trees. We found signifi- cant differences in arthropod community composition between high and low pinyon mortality environments, despite no differences in arthropod abundance or richness. Overall, 22% (51 taxa) of the arthropod community were identified as being indicators of either high or low mortality. Our study corroborates other research indicating that arthropods are responsive to even moderate disturbance events leading to changes in the environment. These arthropod responses can be explained in part due to the increase in woody debris and reduced canopy cover created by tree mortality. RESUMEN.—Documentamos el efecto de la mortalidad causada por la sequía del pino piñonero (Pinus edulis Engelm.) sobre comunidades de artrópodos subterráneos. Utilizamos tres variantes en el microhábitat de los artrópodos incrementando la radiación solar, desechos de madera muerta y vegetación baja.