Marine Genomics Meets Ecology: Diversity and Divergence in South
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ENL 12.Pdf (2.8
, . THE ECHINODERMS NEWSLETTER •I No. 12. Ju1Yt 1982 JIIH B 3 35 PM'63 Distributed by the Department of Invertebrate Zoology, 1 .., ,~ National Museum of Natural History MAl L S E i~VI C L .:;, Smithsonian Institution CEl-iTER Washington, D. C.t U. S. A. " Editor: .fohn M. La'tlTrence ~epartment of Biology Vniversity of South Florida tampat Florida 33620 U. S. A. / / Assistants: Chang-Po Chen, Walter Diehl, Adam Marsh, James McClintockt Stephen Watts ) The Echi~oderms Newsletter was founded in 1968 by David L. Pawson and Maureen E. Downey o~ th~ Smithsonian Institution. They were responsible for the preVious eleven i~sues. Workers with echinoderms, and indeed those who work with fl1 invertebfates, owe them a tremendous debt for the contribution which the #ewsletter has made/to the field. c' "., The neWs~etter generally contains information concerning meetings and conferences and publtcations of interest to echinoderm biologists, titles of theses a,d "-disserta~ions on echinoderms, and research interests and addresses of echlnoderm biologists. To insure inclU$~on~~ journal articles in the newsletter, a~thors are -.requeste4 to send titles or"reprints to the editor. }, ..,.. / SuggestiOns and request,s'from individuals can be addressed to the entire !chinoderm biology ~ommunity th~ough the newsletter. /'~. ," The news~etter ,;f:'s'~~otintendedto be a part of the scientific literature,' and should n~t be/c:ited, abstracted, or reprinted as a published document. •..., " ---- ..•..,->.--------------~--------------------." ./ .' -'. ~:-/.;, Internat+onal Echinoderms Conference--Tampa Bay (1981) /', '.--:. The inte{nati6~al echinoderms conferences began with the Smithsonian meet~ng In 1972. At that conference, participants voted to have subsequent internat!ona1 /lIleetingsevet'! three years, with the site to be changed.between the Ap1eri~as, Europe? fil-ndjtheEast. -

Oceanography and Marine Biology an Annual Review Volume 56
Oceanography and Marine Biology An Annual Review Volume 56 S.J. Hawkins, A.J. Evans, A.C. Dale, L.B. Firth & I.P. Smith First Published 2018 ISBN 978-1-138-31862-5 (hbk) ISBN 978-0-429-45445-5 (ebk) Chapter 5 Impacts and Environmental Risks of Oil Spills on Marine Invertebrates, Algae and Seagrass: A Global Review from an Australian Perspective John K. Keesing, Adam Gartner, Mark Westera, Graham J. Edgar, Joanne Myers, Nick J. Hardman-Mountford & Mark Bailey (CC BY-NC-ND 4.0) Oceanography and Marine Biology: An Annual Review, 2018, 56, 2-61 © S. J. Hawkins, A. J. Evans, A. C. Dale, L. B. Firth, and I. P. Smith, Editors Taylor & Francis IMPACTS AND ENVIRONMENTAL RISKS OF OIL SPILLS ON MARINE INVERTEBRATES, ALGAE AND SEAGRASS: A GLOBAL REVIEW FROM AN AUSTRALIAN PERSPECTIVE JOHN K. KEESING1,2*, ADAM GARTNER3, MARK WESTERA3, GRAHAM J. EDGAR4,5, JOANNE MYERS1, NICK J. HARDMAN-MOUNTFORD1,2 & MARK BAILEY3 1CSIRO Oceans and Atmosphere, Indian Ocean Marine Research Centre, M097, 35 Stirling Highway, Crawley, 6009, Australia 2University of Western Australia Oceans Institute, Indian Ocean Marine Research Centre, M097, 35 Stirling Highway, Crawley, 6009, Australia 3BMT Pty Ltd, PO Box 462, Wembley, 6913, Australia 4Aquenal Pty Ltd, 244 Summerleas Rd, Kingston, 7050, Australia 5Institute for Marine and Antarctic Studies, University of Tasmania, Private Bag 49, Hobart, 7001, Australia *Corresponding author: John K. Keesing e-mail: [email protected] Abstract Marine invertebrates and macrophytes are sensitive to the toxic effects of oil. Depending on the intensity, duration and circumstances of the exposure, they can suffer high levels of initial mortality together with prolonged sublethal effects that can act at individual, population and community levels. -

What Can We Learn from Confusing Olivella Columellaris and O
Biota Neotrop., vol. 12, no. 2 What can we learn from confusing Olivella columellaris and O. semistriata (Olivellidae, Gastropoda), two key species in panamic sandy beach ecosystems? Alison I. Troost1, Samantha D. Rupert1, Ariel Z. Cyrus1,. Frank V Paladino1,2, Benjamin F. Dattilo3 & Winfried S. Peters1,2,4 1Department of Biology, Indiana/Purdue University Fort Wayne, 2101 East Coliseum Boulevard, Fort Wayne, IN 46805‑1499, USA 2Goldring Marine Biology Station, Playa Grande, Santa Cruz, Guanacaste, Costa Rica 3Department of Geosciences, Indiana/Purdue University Fort Wayne, 2101 East Coliseum Boulevard, Fort Wayne, IN 46805‑1499, USA 4Corresponding author: Winfried S. Peters, e‑mail: [email protected] TROOST, A.I., RUPERT, S.D., CYRUS, A.Z., PALADINO, F.V., DATTILO, B.F. & PETERS, W.S. What can we learn from confusing Olivella columellaris and O. semistriata (Olivellidae, Gastropoda), two key species in panamic sandy beach ecosystems? Biota Neotrop. 12(2): http://www.biotaneotropica.org.br/v12n2/ en/abstract?article+bn02112022012 Abstract: Olivella columellaris (Sowerby 1825) and O. semistriata (Gray 1839) are suspension‑feeding, swash‑surfing snails on tropical sandy beaches of the east Pacific. While they often are the numerically dominant macrofaunal element in their habitats, their biology is poorly understood; the two species actually have been confused in all of the few publications that address their ecology. Frequent misidentifications in publications and collections contributed also to an overestimation of the geographic overlap of the two species. To provide a sound taxonomic basis for further functional, ecological, and evolutionary investigations, we evaluated the validity of diagnostic traits in wild populations and museum collections, and defined workable identification criteria. -

The Sea Stars (Echinodermata: Asteroidea): Their Biology, Ecology, Evolution and Utilization OPEN ACCESS
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/328063815 The Sea Stars (Echinodermata: Asteroidea): Their Biology, Ecology, Evolution and Utilization OPEN ACCESS Article · January 2018 CITATIONS READS 0 6 5 authors, including: Ferdinard Olisa Megwalu World Fisheries University @Pukyong National University (wfu.pknu.ackr) 3 PUBLICATIONS 0 CITATIONS SEE PROFILE Some of the authors of this publication are also working on these related projects: Population Dynamics. View project All content following this page was uploaded by Ferdinard Olisa Megwalu on 04 October 2018. The user has requested enhancement of the downloaded file. Review Article Published: 17 Sep, 2018 SF Journal of Biotechnology and Biomedical Engineering The Sea Stars (Echinodermata: Asteroidea): Their Biology, Ecology, Evolution and Utilization Rahman MA1*, Molla MHR1, Megwalu FO1, Asare OE1, Tchoundi A1, Shaikh MM1 and Jahan B2 1World Fisheries University Pilot Programme, Pukyong National University (PKNU), Nam-gu, Busan, Korea 2Biotechnology and Genetic Engineering Discipline, Khulna University, Khulna, Bangladesh Abstract The Sea stars (Asteroidea: Echinodermata) are comprising of a large and diverse groups of sessile marine invertebrates having seven extant orders such as Brisingida, Forcipulatida, Notomyotida, Paxillosida, Spinulosida, Valvatida and Velatida and two extinct one such as Calliasterellidae and Trichasteropsida. Around 1,500 living species of starfish occur on the seabed in all the world's oceans, from the tropics to subzero polar waters. They are found from the intertidal zone down to abyssal depths, 6,000m below the surface. Starfish typically have a central disc and five arms, though some species have a larger number of arms. The aboral or upper surface may be smooth, granular or spiny, and is covered with overlapping plates. -

A Systematic Revision of the Asterinid Genus Aquilonastra O'loughlin
Memoirs of Museum Victoria 63(2): 257–287 (2006) ISSN 1447-2546 (Print) 1447-2554 (On-line) http://www.museum.vic.gov.au/memoirs/index.asp A systematic revision of the asterinid genus Aquilonastra OʼLoughlin, 2004 (Echinodermata: Asteroidea) P. M ARK OʼLOUGHLIN1 AND FRANCIS W.E. ROWE2 1Honorary Associate, Marine Biology Section, Museum Victoria, GPO Box 666, Melbourne, Vic. 3001, Australia ([email protected]) 2Research Associate, Australian Museum, Sydney, NSW, Australia ([email protected]). Private address: Beechcroft, Norwich Road, Scole, Diss, Norfolk, IP21 4DY, U.K. Abstract OʼLoughlin, P. Mark and Rowe, Francis W.E. A systematic revision of the asterinid genus Aquilonastra OʼLoughlin, 2004 (Echinodermata: Asteroidea). Memoirs of Museum Victoria 63(2): 257–287. The Indo-west Pacifi c Aquilonastra OʼLoughlin is reviewed. Eleven species are retained in Aquilonastra: A. anomala (H.L. Clark); A. batheri (Goto); A. burtonii (Gray); A. cepheus (Müller and Troschel); A. corallicola (Marsh); A. coronata (Martens); A. iranica (Mortensen); A. limboonkengi (Smith); A. minor (Hayashi); A. rosea (H.L. Clark); A. scobinata (Livingstone). Asterina lorioli Koehler is reassigned to Aquilonastra. Thirteen new species are described: A. byrneae; A. colemani; A. conandae; A. doranae; A. halseyae; A. marshae; A. moosleitneri; A. oharai; A. richmondi; A. rowleyi; A. samyni; A. watersi; A. yairi. The four subspecies of Asterina coronata Martens are junior synonyms: Asterina coronata cristata Fisher; Asterina coronata euerces Fisher; Asterina coronata fascicularis Fisher; Asterina coronata forma japonica Hayashi. The 13 fi ssiparous Red Sea specimens described by Perrier as Asteriscus wega are the syntypes. Asteriscus wega Perrier is a junior synonym of Asterina burtonii Gray. -

Parks Victoria Technical Series No
Deakin Research Online This is the published version: Barton, Jan, Pope, Adam and Howe, Steffan 2012, Marine protected areas of the Flinders and Twofold Shelf bioregions Parks Victoria, Melbourne, Vic. Available from Deakin Research Online: http://hdl.handle.net/10536/DRO/DU:30047221 Reproduced with the kind permission of the copyright owner. Copyright: 2012, Parks Victoria. Parks Victoria Technical Paper Series No. 79 Marine Natural Values Study (Vol 2) Marine Protected Areas of the Flinders and Twofold Shelf Bioregions Jan Barton, Adam Pope and Steffan Howe* School of Life & Environmental Sciences Deakin University *Parks Victoria August 2012 Parks Victoria Technical Series No. 79 Flinders and Twofold Shelf Bioregions Marine Natural Values Study EXECUTIVE SUMMARY Along Victoria’s coastline there are 30 Marine Protected Areas (MPAs) that have been established to protect the state’s significant marine environmental and cultural values. These MPAs include 13 Marine National Parks (MNPs), 11 Marine Sanctuaries (MSs), 3 Marine and Coastal Parks, 2 Marine Parks, and a Marine Reserve, and together these account for 11.7% of the Victorian marine environment. The highly protected Marine National Park System, which is made up of the MNPs and MSs, covers 5.3% of Victorian waters and was proclaimed in November 2002. This system has been designed to be representative of the diversity of Victoria’s marine environment and aims to conserve and protect ecological processes, habitats, and associated flora and fauna. The Marine National Park System is spread across Victoria’s five marine bioregions with multiple MNPs and MSs in each bioregion, with the exception of Flinders bioregion which has one MNP. -

2006-2007 Intertidal Reef Biodiversity on Kangaroo
2006-2007 Kangaroo Island Natural Resources Management Board INTERTIDAL REEF BIODIVERSITY Intertidal Reef Biodiversity on Kangaroo Island – 2007 ON KANGAROO ISLAND 1 INTERTIDAL REEF BIODIVERSITY ON KANGAROO ISLAND Oceans of Blue: Coast, Estuarine and Marine Monitoring Program A report prepared for the Kangaroo Island Natural Resources Management Board by Kirsten Benkendorff Martine Kinloch Daniel Brock June 2007 2006-2007 Kangaroo Island Natural Resources Management Board Intertidal Reef Biodiversity on Kangaroo Island – 2007 2 Oceans of Blue The views expressed and the conclusions reached in this report are those of the author and not necessarily those of persons consulted. The Kangaroo Island Natural Resources Management Board shall not be responsible in any way whatsoever to any person who relies in whole or in part on the contents of this report. Project Officer Contact Details Martine Kinloch Coast and Marine Program Manager Kangaroo Island Natural Resources Management Board PO Box 665 Kingscote SA 5223 Phone: (08) 8553 4980 Fax: (08) 8553 0122 Email: [email protected] Kangaroo Island Natural Resources Management Board Contact Details Jeanette Gellard General Manager PO Box 665 Kingscote SA 5223 Phone: (08) 8553 0111 Fax: (08) 8553 0122 Email: [email protected] © Kangaroo Island Natural Resources Management Board This document may be reproduced in whole or part for the purpose of study or training, subject to the inclusion of an acknowledgment of the source and to its not being used for commercial purposes or sale. Reproduction for purposes other than those given above requires the prior written permission of the Kangaroo Island Natural Resources Management Board. -

Zoology Honours 2009: Research Projects
Zoology Honours 2009: Research Projects Below are a number of research projects suggested by Animal Biology staff members. This is not the definitive list, and students are encouraged to approach appropriate academic staff within the School of Animal Biology if they have ideas for a research project, or if they want to discuss the possibility of a project within a particular subject area. Staff interests and contact links can be found on the web site [http://www.animals.uwa.edu.au/home/research]. Interested students should contact the Zoology Hons Coordinator [[email protected]] for more details. Conservation genetics: measuring genetic mixing in a translocated population of marine snail Jason Kennington & Mike Johnson [contact [email protected]] Translocation is a management tool that is often used to combat the loss of genetic diversity within small and fragmented populations of rare species (Allendorf & Luikart 2006; Frankham et al. 2002). It involves the movement of individuals between populations with the aim of increasing genetic variation within populations by artificially enhancing gene flow (Storfer 1999; Frankham et al. 2002). In support of this concept, several studies have shown that genetic diversity within small and inbred populations can be restored by natural migration and intentional translocation programs (see Frankham 2005). However, relatively few studies have undertaken post-release monitoring of translocated populations to see if the variation introduced by translocation is maintained over many generations. The aim of this project is to examine the extent of genetic mixing in artificial hybrid populations of the intertidal snail Bembicium vittatum, established in 1993 (Parsons 1997). -

Spatial Distribution of Echinoderms in Littoral Area of Ambon Island, Eastern Indonesia
BIODIVERSITAS ISSN: 1412-033X Volume 19, Number 5, September 2018 E-ISSN: 2085-4722 Pages: 1919-1925 DOI: 10.13057/biodiv/d190544 Spatial distribution of echinoderms in littoral area of Ambon Island, Eastern Indonesia ANA SETYASTUTI1,♥, WAHYU PURBIANTORO2, HADIYANTO1 1Research Center for Oceanography, Indonesian Institute of Sciences. Jl. Pasir Putih Raya No.1, North Jakarta 14430, Jakarta, Indonesia. Tel.: +62-21- 64713850, email: [email protected] 2Research Center for Deep Sea, Indonesian Institute of Sciences. Jl. Pasir Putih Raya No.1, North Jakarta 14430, Jakarta, Indonesia Manuscript received: 17 July 2018. Revision accepted: 27 September 2018. Abstract. Setyastuti A, Purbiantoro W, Hadiyanto. 2018. Spatial distribution of echinoderms in littoral area of Ambon Island, Eastern Indonesia. Biodiversitas 19: 1919-1925. Echinoderms samples were collected in Ambon Island as part of the marine resource inventories designed to study the biodiversity of Maluku Archipelago. The purpose of this works was to investigate the abundance and genus richness of echinoderms within the five sampling sites using single transect orthogonal to the coast consisted of 10-quadrat plot of 5x5m, the distance between plots was 10m at each site. Furthermore, this study also aimed to understand the relation of substrate type and community composition within those five sites. The substrate was classified into four types (sea-grass, macro-algae, sand, rocks and/or dead coral) noted by presence/absence mark at each plot. The differences in total abundance and genus richness among those sites were analyzed using an Analysis of Deviance following a Generalized Linear Models (GLMs) fitted by Poisson and log link. However, in term of genus composition, the data were visualized using a Principal Coordination Analysis (PCoA) plots calculated from Bray-Curtis dissimilarity based on the log (x+1) transformed abundance data. -
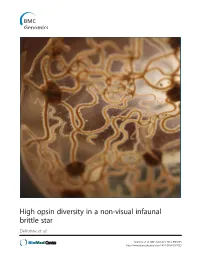
Amphiura Filiformis, We First Highlighted a Blue-Green Light Sensitivity Using a Behavioural Approach
High opsin diversity in a non-visual infaunal brittle star Delroisse et al. Delroisse et al. BMC Genomics 2014, 15:1035 http://www.biomedcentral.com/1471-2164/15/1035 Delroisse et al. BMC Genomics 2014, 15:1035 http://www.biomedcentral.com/1471-2164/15/1035 RESEARCH ARTICLE Open Access High opsin diversity in a non-visual infaunal brittle star Jérôme Delroisse1*, Esther Ullrich-Lüter2, Olga Ortega-Martinez3, Sam Dupont3, Maria-Ina Arnone4, Jérôme Mallefet5 and Patrick Flammang1 Abstract Background: In metazoans, opsins are photosensitive proteins involved in both vision and non-visual photoreception. Echinoderms have no well-defined eyes but several opsin genes were found in the purple sea urchin (Strongylocentrotus purpuratus) genome. Molecular data are lacking for other echinoderm classes although many species are known to be light sensitive. Results: In this study focused on the European brittle star Amphiura filiformis, we first highlighted a blue-green light sensitivity using a behavioural approach. We then identified 13 new putative opsin genes against eight bona fide opsin genes in the genome of S. purpuratus. Six opsins were included in the rhabdomeric opsin group (r-opsins). In addition, one putative ciliary opsin (c-opsin), showing high similarity with the c-opsin of S. purpuratus (Sp-opsin 1), one Go opsin similar to Sp-opsins 3.1 and 3.2, two basal-branch opsins similar to Sp-opsins 2 and 5, and two neuropsins similar to Sp-opsin 8, were identified. Finally, two sequences from one putative RGR opsin similar to Sp-opsin 7 were also detected. Adult arm transcriptome analysis pinpointed opsin mRNAs corresponding to one r-opsin, one neuropsin and the homologue of Sp-opsin 2. -

Population Ecology of the Intertidal Sea Star Asterina Burtoni
Qatar Univ. Sci. J. (1995), 15 (1): 163 • 171 POPULATION ECOLOGY OF THE INTERTIDAL SEA STAR ASTERINA BURTON! GRAY (ASREROIDEA: ECHINODERMATA) IN THE WAKRAH BAY, SOUTH EAST OF QATAR By F. EL. SOLIMAN Department of Zoology, Faculty Of Science, University of Qatar, Doha, Qatar (~l:S"_,.!-~: ~' .. -~1} ":?'.r.- «~J1J..t! L:J_,.:-1» .r--:JI ~ ;~ 4. )ai 4JJ,) J..,..!t ~~ o..,s'}I ~ ;~IJ ..UI Ula:..o ~ ~ • !)L..J..., ..\.....JI .. • ~ .... ~ ~Y.Jfi t.:,._r-1» .r--:ll ~ ~~ ~.j~l ~.rJIJ ~J,:liJ ~I,:JI t...l;,) ~11.11& J f erJII.lla ~l 4-.I;.UI ~Jt .liJ, )ai t)J,) J~ ~~ o_?_,ll ~ ;-*IJ .U.I ~ «~.jlr. ~1 J ~I.,_.!.P t}.J-A erJII.lla ~ J~ •;~I ~t L.S ~.r _r.o JS' J Y, Y ~,)..\P 4i~ ~..t! ~lzll ~Jl ~. i~l ~ J>-b ~ taJa.a ~I USI_,Jall ~.1>l ~~~I ~J~ ~I ~jJ) ~I ;I..L:I: ~\.J:-1 ~j_,JIJ t!;.UI J_,k 'I" 4 41·..:... ~L....y ~')U f1..u;.:....4 +\'I"~ USI_,k ~) Ji'il ~ lf! ~..t! i~l ~t ~~I~ ~Y:-1 ~j_,JIJ (~L.!.>~I ~f ~ J i~l ~1 ~I ~I ~I USI_,Jall ~..L>t ~11iW~4 +tJ +Y"J +yJ ,:r i.rAJI J~ ~WI ~'-~4 ~I ~I USI}alJ ;~'it ~Ua...._,:.- ~;..U L.S. \\\Y" .r'""t ~~ ~J '""'" ,...,pt Key Words: Asterina burtoni, Population ecology, Age class, Evacuated body, Gonads, Pyloric caeca ABSTRACT The abundance, distribution and population age structure of the sea star Asterina burtoni Gray were investigated in the intertidal area of the Wakrah Bay, South East Qatar Peninsula from October, 1993 to January, 1994. -

Metabarcoding Analysis on European Coastal Samples Reveals New
www.nature.com/scientificreports OPEN Metabarcoding analysis on European coastal samples reveals new molecular metazoan diversity Received: 8 November 2017 David López-Escardó1, Jordi Paps2, Colomban de Vargas3,4, Ramon Massana5, Accepted: 5 June 2018 Iñaki Ruiz-Trillo 1,6,7 & Javier del Campo1,5 Published: xx xx xxxx Although animals are among the best studied organisms, we still lack a full description of their diversity, especially for microscopic taxa. This is partly due to the time-consuming and costly nature of surveying animal diversity through morphological and molecular studies of individual taxa. A powerful alternative is the use of high-throughput environmental sequencing, providing molecular data from all organisms sampled. We here address the unknown diversity of animal phyla in marine environments using an extensive dataset designed to assess eukaryotic ribosomal diversity among European coastal locations. A multi-phylum assessment of marine animal diversity that includes water column and sediments, oxic and anoxic environments, and both DNA and RNA templates, revealed a high percentage of novel 18S rRNA sequences in most phyla, suggesting that marine environments have not yet been fully sampled at a molecular level. This novelty is especially high among Platyhelminthes, Acoelomorpha, and Nematoda, which are well studied from a morphological perspective and abundant in benthic environments. We also identifed, based on molecular data, a potentially novel group of widespread tunicates. Moreover, we recovered a high number of reads for Ctenophora and Cnidaria in the smaller fractions suggesting their gametes might play a greater ecological role than previously suspected. Te animal kingdom is one of the best-studied branches of the tree of life1, with more than 1.5 million species described in around 35 diferent phyla2.