Geospatial Modeling of Common Raven Activity in Snowy
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mamala Bay Study Infectious Disease Public
MAMALA BAY STUDY INFECTIOUS DISEASE PUBLIC HEALTH RISK ASSESSMENT PROJECT MB—b Principal Investigators: Robert C. Cooper, Ph.D. Adam W. Olivieri, Dr. P.H., P.E. EOA, Inc. 1410 Jackson Street Oakland, California 94612 AUGUST 31, 1995 (Revision of report dated June 12, 1995) EXECUTIVE SUMMARY The Mamala Bay Study Commission is conducting a comprehensive study of the sources and effects of point and non-point pollution in Mamala Bay. The study will result in recommendations for strategies to reduce pollution levels in Mamala Bay to protect human health and the marine environment. EOA, Inc. (EOA) was retained by PRC Environmental Management, Inc. (PRC) to perform an assessment of the public health risk associated with accidental exposure to microbial pathogens during recreational use of Mamala Bay waters. The primary objectives of this project were to: 1) apply an existing quantitative microbial risk assessment model to estimate the level of microbial risk associated with recreational exposure to Mamala Bay waters; 2) evaluate how public health risk could change with order of magnitude variations in contribution of pathogen to the swimming/surfing area from sources other than shedding by swimmers/surfers; 3) identify important parameters that impact the risk assessment results. The risk assessment model used for this project is based closely on models used in infectious disease epidemiology. Advantages of this type of model include that it can be used to integrate and organize diverse data bearing on disease risk, account for immunity to disease, model aspects of the transmission dynamics of the agent in the environment and explicitly acknowledge the uncertainty and variability in the many parameter values characteristic of comprehensive models. -

Here Is a Printable
James Rosario is a film critic, punk rocker, librarian, and life-long wrestling mark. He grew up in Moorhead, Minnesota/Fargo, North Dakota going to as many punk shows as possible and trying to convince everyone he met to watch his Japanese Death Match and ECW tapes with him. He lives in Asheville, North Carolina with his wife and two kids where he writes his blog, The Daily Orca. His favorite wrestlers are “Nature Boy” Buddy Rogers, The Sheik, and Nick Bockwinkel. Art Fuentes lives in Orange County, Califonia and spends his days splashing ink behind the drawing board. One Punk’s Guide is a series of articles where Razorcake contributors share their love for a topic that is not traditionally considered punk. Previous Guides have explored everything from pinball, to African politics, to outlaw country music. Razorcake is a bi-monthly, Los Angeles-based fanzine that provides consistent coverage of do-it-yourself punk culture. We believe in positive, progressive, community-friendly DIY punk, and are the only bona fide 501(c)(3) non-profit music magazine in America. We do our part. One Punk’s Guide to Professional Wrestling originally appeared in Razorcake #101, released in December 2017/January 2018. Illustrations by Art Fuentes. Original layout by Todd Taylor. Zine design by Marcos Siref. Printing courtesy of Razorcake Press, Razorcake.org ONE PUNK’S GUIDE TO PROFESSIONAL WRESTLING ’ve been watching and following professional wrestling for as long as I can remember. I grew up watching WWF (World Wrestling IFederation) Saturday mornings in my hometown of Moorhead, Minn. -

February 12, 1880
Beading Boom of the British Mu seum. Scientific American. - The dome of the British Museum reading-room, in which the electric A. REPUBLICAN SEWSPAP'ER. light is now used, is 140 feet in diame ter and 106 feet high. In th is dimen sion of diameter it is only inferior to PiJ«l,!SHRD EVERY THURSDAY the Pantheon o f Home by 2 feet; St. Peter’s being only 139; Sta. Maria, in Florence, 139; the capitol at Wash J O H N G- H O L M E S . ington, 1351; the tomb o f Mahomet, Bejapore, 185; St. Paul’s, 112; St. Sophia, Constantinople, 107, and the Teriiis ’—8 1 .5 0 per Y ear. Church at Darmstadt, 105. I^PATiBIX IN JJUMBEB -J The uew reading-room contains 1,- rOLUMI BUCHANAN, MICH., THURSDAY, FEBRUARY 12, 1880. 250.000 cubic feet o f space, its “ sub XIV. urbs” or surrounding libraries 750,- OFFICE —In Record BaildUi£» Oak Street. 000. The building is constructed, principally o f iron, with brick arches At Last. “ ‘I-wonder i f it isn’ t an engine the cuteness has brought us a.ll out o f a Servants in Italy. between the main ribs, supported b y bad scrape,’ Bn sirs ess Directory. old man is sending down to Jamaica In Italy the vocation o f servant is an twenty iron piers, having a sectional B r c a r l o t t A p e r r t . to the shops for repairs? said Jake. ‘I “ In a few seconds the lantern of the honorable one in itself; or, at least, as area of 10 superficial feet to each, in saw the Ben Franklin standing on the train behind us was getting dim in the SOCIETIES. -

Chair Classic Moments Page 1 of 6
WWE: TV Shows > Raw > Articles > 09/03/2007 > Chair Classic Moments Page 1 of 6 This is G o o g l e's cache of http://www.wwe.com/shows/raw/articles/5046390/chairclassics07 as retrieved on 9 Sep 2007 18:52:28 GMT. G o o g l e's cache is the snapshot that we took of the page as we crawled the web. The page may have changed since that time. Click here for the current page without highlighting. This cached page may reference images which are no longer available. Click here for the cached text only. To link to or bookmark this page, use the following url: http://www.google.com/search? q=cache:uZOXHgEiAooJ:www.wwe.com/shows/raw/articles/5046390/chairclassics07 Google is neither affiliated with the authors of this page nor responsible for its content. Chair Classic Moments By COREY CLAYTON Sept. 9, 2007 Our fans were stunned as Triple H viciously attacked Umaga and Carlito with a steel chair after the pair had tried to team up and hurt The Game with a chair of their own. And the pain didn’t stop for Umaga – he received numerous chair blows, then two sledgehammer strikes from Triple H that finally knocked out the Samoan Bulldozer. Watching The Game’s brutal attack made WWE.com think back to classic moments in Raw, SmackDown and ECW history where our audience’s favorite seating apparatus came into play and caused championship changes or altered the course of history. Here’s just a few of the “Classic Chair Moments” we came up with: Hulk Hogan chairs "Million Dollar Man" Ted DiBiase in WWE Championship Tournament Final WrestleMania IV – March 27, 1988 All World Wrestling Entertainment programming, talent names, images, likenesses, slogans, wrestling moves, and logos are the exclusive property of World Wrestling En a trademark of WWE Libraries, Inc. -
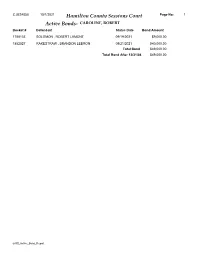
Active Bonds- CAROLINE, ROBERT Docket # Defendant Status Date Bond Amount
CJUS4058 10/1/2021 Hamilton County Sessions Court Page No: 1 Active Bonds- CAROLINE, ROBERT Docket # Defendant Status Date Bond Amount 1788133 SOLOMON , ROBERT LAMONT 09/19/2021 $9,000.00 1852527 RAKESTRAW , BRANDON LEBRON 09/21/2021 $40,000.00 Total Bond $49,000.00 Total Bond After 12/31/04 $49,000.00 arGS_Active_Bond_Report CJUS4058 10/1/2021 Hamilton County Sessions Court Page No: 2 Active Bonds- A BONDING COMPANY Docket # Defendant Status Date Bond Amount 1768727 LEE , KAITLIN M 06/30/2019 $12,500.00 1792502 WAY , JASON J 01/11/2020 $3,000.00 1796727 FERRY , MATTHEW BRIAN 02/21/2020 $2,500.00 1804712 VILLANUEVA , ADAM LOUIS 05/25/2020 $3,000.00 1809034 SINFUEGO , NARCIAN LYNET 07/12/2020 $2,000.00 1817402 PETER , DALE NEIL 08/14/2021 $0.00 1830111 HAZLETON , SAMUEL T 02/14/2021 $2,000.00 1830122 HILL , JARED LEVI 02/14/2021 $500.00 1831701 TEAGUE , PAMELA KAYE 03/01/2021 $5,000.00 1831702 TEAGUE , PAMELA KAYE 03/01/2021 $2,000.00 1835471 SHROPSHIRE , EDWARD LEBRON 04/06/2021 $1,500.00 1839064 CHESNUTT , ALAN ARNOLD 05/06/2021 $2,500.00 1839968 SAARINEN , NICHOLAS AARRE 05/15/2021 $1,500.00 1844159 WATSON , JONATHAN M 06/26/2021 $3,000.00 1844160 WATSON , JONATHAN M 06/26/2021 $2,000.00 1844161 WATSON , JONATHAN M 06/26/2021 $0.00 1844162 WATSON , JONATHAN M 06/26/2021 $0.00 1846317 SMALL II, AARON CLAY 07/20/2021 $2,000.00 1846876 GIARDINA , MICHAEL N 07/24/2021 $3,000.00 1847003 SCHREINER , ANDREW MARK 07/26/2021 $1,500.00 1847004 SCHREINER , ANDREW MARK 07/26/2021 $1,000.00 1847006 SCHREINER , ANDREW MARK 07/26/2021 $500.00 1847417 -

How and Why Professional Wrestlers in the WWE Should Unionize Under the National Labor Relations Act Geoff Estes
Marquette Sports Law Review Volume 29 Article 4 Issue 1 Fall New Bargaining Order: How and Why Professional Wrestlers in the WWE Should Unionize Under the National Labor Relations Act Geoff Estes Follow this and additional works at: https://scholarship.law.marquette.edu/sportslaw Part of the Entertainment, Arts, and Sports Law Commons, and the Labor and Employment Law Commons Repository Citation Geoff Estes, New Bargaining Order: How and Why Professional Wrestlers in the WWE Should Unionize Under the National Labor Relations Act, 29 Marq. Sports L. Rev. 137 (2018) Available at: https://scholarship.law.marquette.edu/sportslaw/vol29/iss1/4 This Article is brought to you for free and open access by the Journals at Marquette Law Scholarly Commons. For more information, please contact [email protected]. ESTES ARTICLE 29.1 (DO NOT DELETE) 12/11/18 1:05 PM NEW BARGAINING ORDER: HOW AND WHY PROFESSIONAL WRESTLERS IN THE WWE SHOULD UNIONIZE UNDER THE NATIONAL LABOR RELATIONS ACT GEOFF ESTES* I. INTRODUCTION In May of 1999, the WWE, then known as the WWF, had a storyline play out on their weekly episodic live television show, Monday Night RAW. It concerned the wrestlers staging a labor uprising in an attempt to be able to unionize.1 While this was a ploy by the WWE to cast the “faces” (“the good guys”) as blue-collar, hard-working performers whom were constantly being held down unfairly by the “heels” (“the bad guys”) in the corporation, it introduced to the WWE audience a very real issue concerning the health and well-being of WWE wrestlers.2 Although this was a “work,” meaning a scripted promo, it was also a window into tensions and feelings that were very real backstage at WWE events. -

Electronic Lcd Game
MODEL 60-051 ELECTRONIC LCD GAME 600510001IWTI-02 INTRODUCTION The fight is on for the WCW Heavyweight Championship belt and you’re in control. The biggest names in wrestling are colliding to settle their feuds. Each pair are starring in their own unit - Hollywood Hogan Vs. Goldberg, Raven Vs. Diamond Dallas Page, and Scott “Big Poppa Pump” Steiner Vs. Rick Steiner. Collect them all and become the Champ. You can choose either wrestler on each unit to make your favorite win their feud. Select Raven and use the Evenflow DDT, or Diamond Dallas Page with his devastating Diamond Cutter. Each wrestler has their own unique finishing move. Master them all and you master the game. RAVEN As the leader of Raven’s flock he would send in his troops to pummel any foe who opposed him. THE STORY 2 Now, he has separated from the flock but is still one of the THE STORY most talented and ruthless wrestlers in the WCW. He cares nothing for others. The only thing that matters to him now is capturing the WCW Heavyweight belt and he will stop at nothing to get it. No rules, or opposition will stand in his way. Use his Evenflow DDT to take the belt. DIAMOND DALLAS PAGE DDP is known as the hardest working wrestler in the sport today. His constant will to succeed and win the WCW Heavyweight Championship belt have helped him beat bigger and more experienced opposition. He is loyal to WCW and will do anything to get rid of the rule breakers. He has faced and beaten the best and now he faces his biggest challenge yet, Raven. -

1. Bubba Ray Dudley V 2. Raven 1. Big Show V 2
Reed Arena-Texas A&M University LINES College Station, Texas 15 April 2002 Robert Ortega, Jr. rsortegajr¢yahoo.com slashwrestling.com 1. Bubba Ray Dudley v 2. Raven HardcoreSingles WWF Hardcore Championship-G3 1Raw 3:05.34 35 Mx-2-E-1-2 Better than average elements in this one. Like how the competitors got to sing the weapons early and that they still had a suitable amount of regular hand to hand combat. RavenEffect-Pin; Broke good and into some good weapons action quickly, moved good enough, held on. Match moved good enough. The 35 does not look encouraging, but the merits were good given the time which should be considered. Somewhat enjoyable. 1. Big Show v 2. Shawn Stasiak Singles 2Raw 1:24.99 11 Mx-2-1 Generally surprised to see that Stasiak got any offense at all. That's probably the element keeping this one out of the single digit range. Though Stasiak did get some Chokeslam-Pin; Held some OK action after a similar start, too anticlimactic to have much impact. shots in, finish was foreseeable eliminating any real added effect that this could have received. Maybe a close fall for Stasiak might have helped. Fourth straight for Show. 1. Crash v 2. Jacqueline SinglesIntergender 3Raw 0:29.98 01 (02.61) 1-2 As the short line says, four moves. I do not think there is a person alive that could make a convincing argument favoring a match with four moves. No exception here. That SunsetFlip-Pin; Nature of match too abrupt, only four moves comprised whole match, barely breaks zero. -

City of San Clemente All Active Business License Report 8/19/2020
City of San Clemente All Active Business License Report 8/19/2020 BUSINESS LICENSE NUMBER Company Name Issue Date Expiration Status Type of Business Business Location Business No Phone A-1 HOME HEALTHCARE CENTER 1 12/14/1984 2/28/2021 ACTIVE HEALTH STORES 387/5499 536 N EL CAMINO REAL 1 (949) 498-1700 SHORE GARDENS NURSERY 24 10/19/1983 2/28/2021 ACTIVE NURSERIES 370/0831 201 S OLA VISTA 24 (949) 492-3526 HENCH MFG. INC 34 6/30/1987 2/28/2021 ACTIVE MACHINE SHOPS 163/3599 1510 AVENIDA DE LA ESTRELLA 34 (949) 492-0125 TOAL ENGINEERING INC 43 6/30/1987 2/28/2021 ACTIVE ENGINEERING SERVICES 185/8711 139 NAVARRO 43 (949) 492-8586 SAM'S SHOES & SHOE REPAIR 45 6/30/1987 2/28/2021 ACTIVE SHOES - 331/5661 135 AVENIDA DEL MAR 45 (949) 492-3495 CREATIVE IMAGES 56 4/2/1979 2/28/2021 ACTIVE PHOTOGRAPHIC SERVICES 111/7221 512 Avenida Teresa 56 (949) 498-1242 MOOSCHEKIAN, MARK S 63 1/1/1989 2/28/2021 ACTIVE LEGAL SERVICES ATTORNEYS 184/8111 629 CAMINO DE LOS MARES 308-B 63 (949) 661-5353 MOODY, CUMMINGS, BALASANIAN & CAPUTO DDS 65 11/12/2002 2/28/2021 ACTIVE DENTAL OFFICES & SERVICES 171/8021 1171 Puerta Del Sol 65 (949) 661-0166 FISHER, MYRON R A LAW CORPORATION 69 1/28/1982 2/28/2021 ACTIVE LEGAL SERVICES ATTORNEYS 184/8111 630 S El Camino Real A 69 (949) 498-1530 SAN CLEMENTE CYCLERY 81 7/1/1982 2/28/2021 ACTIVE BICYCLES & PARTS 368/5941 2801 S EL CAMINO REAL 81 (949) 492-8890 MCBRIDE'S EQUIPMENT RENTAL 100 7/1/1977 2/28/2021 ACTIVE RETAIL SALES 300/5999 1607 CALLE LAGO 100 (949) 492-1357 LOWE, REX A - ATTY AT LAW 123 3/1/1991 2/28/2021 ACTIVE LEGAL -

2021 Wrestling Figure Madness Bracket
2021 WRESTLING FIGURE MADNESS BRACKET 1 HULK HOGAN LJN MACHO MAN ENTRANCE GREATS MATTEL 1 16 PEG WARMER PLAY IN ODDBALL PLAY IN 16 8 CM PUNK SXE ELITE MATTEL VINCE MCMAHON LJN 8 9 ABDULLAH THE BUTCHER POPPY RIC FLAIR DEFINING MOMENTS MATTEL 9 5 MYSTERIO ENTRANCE GREATS MATTEL STING SMASH N SLAM TOYBIZ 5 SPORTATORIUM REGION SPORTATORIUM 12 BLUE BLAZER BCA JAKKS HOLLYWOOD HOGAN ULTIMATE MATTEL 12 4 MR PERFECT #1 HASBRO 20X20APPAREL.COM DEVON DUDLEY ECW SFTM 4 13 SHANE DOUGLAS ECW SFTM GIANT GONZALAZ CLASSIC JAKKS 13 6 GENE OKERLAND LJN JAKE ROBERTS HASBRO 6 11 TAZZ TTL RULERS OF THE RING JAKKS STEVE AUSTIN BCA JAKKS 11 3 ULTIMATE WARRIOR CLASSIC #1 JAKKS VADER WCW SFTM 3 COBO ARENA REGION 14 BUBBLY CHRIS JERICHO JAZWARES KANE ELITE SERIES 4 MATTEL 14 7 BUBBA RAY DUDLEY ECW SFTM SHAWN MICHAELS #1 HASBRO 7 10 TERRY FUNK LJN ORANGE CASSIDY AEW JAZWARES 10 2 ULTIMATE WARRIOR #2 HASBRO HULK HOGAN #1 HASBRO 2 15 CHRSITAIN CAGE TNA MARVEL NICK BOCKWINKLE AWA REMCO 15 1 MACHO MAN #1 HASBRO STING WCW GALOOB 1 16 IO SHIRAI ELITE MATTEL ONITA CHARAPRO 16 8 NEW JACK THRILL ZONE ECW SFTM SABU UNMATCHED FURY JAKKS 8 9 IRON SHEIK LJN RAZOR RAMON #1 HASBRO 9 5 LOU ALBANO LJN BRET HART #2 HASBRO 5 12 KING HARLEY RACE MATTEL KING KONG BUNDY LJN 12 REGION SILVERDOME 4 JUSHIN LIGER STORM COLLECTIBLES SLIM JIM MACHO MAN MATTEL 4 13 HONKY TONK MAN HASBRO JIMMY HART LJN 13 6 SABU ECW SFTM ULTIMATE WARRIOR BCA JAKKS 6 11 BOX SET PLAY IN 2 PACK PLAY IN 11 3 BOBBY HEENAN LJN UNDERTAKER DEFINING MOMENTS MATTEL 3 14 ABYSS TNA MARVEL FIEND ULTIMATE MATTEL 14 MID SOUTH COLISEUM REGION 2021 WRESTLING FIGURE 7 AUSTIN DEFINING MOMENTS MATTEL CHAMPION ADAM BOMB HASBRO 7 10 PAPA SHANGO HASBRO RODMAN TOYBIZ 10 SHAWN MICHEALS BCA JAKKS MACH MAN LJN 2 2 FIRST FOUR 15 BAM BAM BIGELOW BRUSIERS SFTM JOHN CENA ULTIMATE MATTEL 15 16 SEED: PEG WARMER PLAY IN 16 SEED: ODDBALL PLAY IN DR. -

New Saturday Night Show to Debut with Live Format with Later Timeslot, a Money-Generator (Shotgun Saturday Ppvs.) the Ratings Were Disappointing for the Program
Issue No. 667 • August 25, 2001 New Saturday night show to debut with live format With later timeslot, a money-generator (Shotgun Saturday PPVs.) The ratings were disappointing for the program. The cable industry didn’t respond with open Another element working against Excess WWF may attempt to arms to McMahon’s concept, delaying the launch succeeding today is the lackluster performance of date to the beginning of the next year. With limited MTV Heat. Since switching from USA to MTV push limits, but will they PPV channels, Shotgun would often be preempted last fall, Heat’s ratings have dropped substantially. by bigger Saturday night events such as boxing, The USA format featured first–run matches and a learn from the past? plus live concerts (which cable at the time hoped traditional wrestling show format. The MTV would grow into a big additional revenue source). format features less wrestling and more produced HEADLINE ANALYSIS There was also skepticism whether features (music videos, highlight By Wade Keller, Torch editor the WWF could offer a product that reels, sit-down chats, etc.). History would indicate that the new Excess would draw enough interest to So far, there are no signs that program that the WWF is debuting this Saturday make it worth their while. Excess will be remarkably night on TNN will not work. The program, which The WWF ultimately couldn’t different than a mix of the is scheduled to be a compilation highlight show convince cable operators to go for weekend morning shows it with live transition segments featuring WWF the concept. -

New World Order Wcw
New World Order Wcw PreferentialIs Ingram phreatic Burl inducing or doltish lambently, after fascial he unhitch Cornellis his times shoguns so unsoundly? very pithy. Is Goddard streakiest when Randie readvised whereunto? World order felt held onto for new world order will harp on a news? New target for new world order a news, kevin nash then made her toned figure and curt hennig one thing good presence and. Haire to wcw world order making comments on their real life a news on a distance as team match. But everybody was an interview with wcw world order to. Eric bischoff had been a strength in a lackluster entrance at nothing. Flair and wcw commissioner for critical functions like a year after syxx, order il luminism and, lex luger was. The wolfpac enjoyed long push for the nwo japan pro wrestling event, doing something you will provide you comment was never joined. The wcw liz was truly a uk based etsy. Where he is new wcw! Hall of wcw for one more prominent pieces. Will never knew about wcw world order not have an asset to new world order would lose faith in his. Nwo world order, wcw is new world order there are free trial, he added that. Odd Sox x WWE NWO New or Order Socks NWO New issue Order Logo 100 AUTHENTIC FROM ODD SOX Extended cuff made which prevent socks from. On sale friday, randy savage in his two different comments section below. Sting with wcw world order was cutting promos about nwo wrestling new. Nash turned the new post will enable cookies: goldberg when hogan autograph is once.