Seagrass Meadows As Seascape Nurseries for Rockfish (Sebastes Spp.)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Emerging Understanding of Seagrass and Kelp As an Ocean Acidification Management Tool in California
EMERGING UNDERSTANDING OF SEAGRASS AND KELP AS AN OCEAN ACIDIFICATION MANAGEMENT TOOL IN CALIFORNIA Developed by a Working Group of the Ocean Protection Council Science Advisory Team and California Ocean Science Trust AUTHORS Nielsen, K., Stachowicz, J., Carter, H., Boyer, K., Bracken, M., Chan, F., Chavez, F., Hovel, K., Kent, M., Nickols, K., Ruesink, J., Tyburczy, J., and Wheeler, S. JANUARY 2018 Contributors Ocean Protection Council Science Advisory Team Working Group The role of the Ocean Protection Council Science Advisory Team (OPC-SAT) is to provide scientific advice to the California Ocean Protection Council. The work of the OPC-SAT is supported by the California Ocean Protection Council and administered by Ocean Science Trust. OPC-SAT working groups bring together experts from within and outside the OPC-SAT with the ability to access and analyze the best available scientific information on a selected topic. Working Group Members Karina J. Nielsen*, San Francisco State University, co-chair John J. Stachowicz*, University of California, Davis, co-chair Katharyn Boyer, San Francisco State University Matthew Bracken, University of California, Irvine Francis Chan, Oregon State University Francisco Chavez*, Monterey Bay Aquarium Research Institute Kevin Hovel, San Diego State University Kerry Nickols, California State University, Northridge Jennifer Ruesink, University of Washington Joe Tyburczy, California Sea Grant Extension * Denotes OPC-SAT Member California Ocean Science Trust The California Ocean Science Trust (OST) is a non-profit organization whose mission is to advance a constructive role for science in decision-making by promoting collaboration and mutual understanding among scientists, citizens, managers, and policymakers working toward sustained, healthy, and productive coastal and ocean ecosystems. -

Coastal Environment and Seaweed-Bed Ecology in Japan
Kuroshio Science 2-1, 15-20, 2008 Coastal Environment and Seaweed-bed Ecology in Japan Kazuo Okuda* Graduate School of Kuroshio Science, Kochi University (2-5-1, Akebono, Kochi 780-8520, Japan) Abstract Seaweed beds are communities consisting of large benthic plants and distributed widely along Japanese coasts. Species constituting seaweed beds in Japan vary depending on localities because of the influence of cold and warm currents. Seaweed beds are important producers along coastal eco- systems in the world, but they have been reduced remarkably in Japan. Not only artificial construc- tions in seashores but also the Phenomenon called “isoyake” have led to the deterioration of coastal environments. To figure out what is going on in nature, Japanese Government began the nationwide survey of seaweed beds. Although trials to recover seaweed communities have been also carrying out, they are not always the solution of subjects. We should think of harmonious coexistence between nature and human being so that problems might not happen. beds and coastal environments; and 5) Towards the sus- Introduction tainable coexistence between nature and human beings Through four billion years of evolution, life on 1. Diversity and distribution of seaweed beds in earth has expanded to almost infinite diversity, with each Japan species interacting with others and molding itself to its habitat until a global ecosystem developed. This diver- Japan has wide range of climate zones from cold sity of life forms is commonly referred to as biodiversity. temperate to subtropical, reflecting its wide geographical Biodiversity is not only crucial to ecosystem balance, but extent from north to south, as well as the influence of also brings great benefit to human lives. -

Blue Carbon Sequestration Along California's Coast
Briefing heldDecember 2020 CCST Expert Briefing Series A Carbon Neutral California One For more details Pager about this briefing: Blue Carbon Sequestration along California’s Coast ccst.us/expert-briefings Select Experts The following experts can advise on Blue Carbon pathways: Joanna Nelson, PhD Founder and Principal LandSea Science [email protected] http://landseascience.com/ Expertise: salt marsh ecology, coastal ecosystem conservation and resilience Lisa Schile-Beers, PhD Senior Associate Silvestrum Climate Associates Figure: Carbon cycles in coastal (blue carbon) habitats (The Watershed Company) Research Associate Smithsonian Environmental Background Research Center • Anthropogenic carbon emissions are a • Blue Carbon refers to carbon stored by [email protected] leading cause of climate change. coastal ecosystems including wetlands, salt Office: (415) 378-2903 marshes, seagrass meadows, and kelp forests. Expertise: wetland and marsh ecology, • California has set an ambitious goal of being carbon cycling, and sea level rise carbon neutral by 2045. • Restoring coastal habitats can increase blue • A combined approach of reducing emissions carbon sequestration and contribute to Melissa Ward, PhD and sequestering carbon – physically state goals. Post-doctoral Researcher removing CO2 from the atmosphere and San Diego State University [email protected] storing it long-term – can help California • Restored coastal habitats also provide many https://melissa-ward.weebly.com reach its goals. other co-benefits. Expertise: carbon storage in seagrass, SEQUESTERING Blue Carbon marsh, and kelp ecosystems in California’s Coastal Ecosystems Lisamarie Per unit area, coastal wetlands, marshes, and Benefits of Blue Carbon Habitats Windham-Myers, PhD eelgrass meadows capture more carbon than 1. Reduced atmospheric CO2 levels Research Ecologist terrestrial habitats such as forests. -

Plymouth Sound and Estuaries SAC: Kelp Forest Condition Assessment 2012
Plymouth Sound and Estuaries SAC: Kelp Forest Condition Assessment 2012. Final report Report Number: ER12-184 Performing Company: Sponsor: Natural England Ecospan Environmental Ltd Framework Agreement No. 22643/04 52 Oreston Road Ecospan Project No: 12-218 Plymouth Devon PL9 7JH Tel: 01752 402238 Email: [email protected] www.ecospan.co.uk Ecospan Environmental Ltd. is registered in England No. 5831900 ISO 9001 Plymouth Sound and Estuaries SAC: Kelp Forest Condition Assessment 2012. Author(s): M D R Field Approved By: M J Hutchings Date of Approval: December 2012 Circulation 1. Gavin Black Natural England 2. Angela gall Natural England 2. Mike Field Ecospan Environmental Ltd ER12-184 Page 1 of 46 Plymouth Sound and Estuaries SAC: Kelp Forest Condition Assessment 2012. Contents 1 EXECUTIVE SUMMARY ..................................................................................................... 3 2 INTRODUCTION ................................................................................................................ 4 3 OBJECTIVES ...................................................................................................................... 5 4 SAMPLING STRATEGY ...................................................................................................... 6 5 METHODS ......................................................................................................................... 8 5.1 Overview ......................................................................................................................... -

CHAPERONE Kelp Vs
4th GRADE Things to do GALLERY Reef Level 1 Level Level 2 Level CHANGING EXHIBIT Tropical Tunnel Tropical GALLERY F I E L D T R I P Tropical CHANGING EXHIBIT Tropical Tunnel Tropical Main Entrance Live Coral Live Tropical Tunnel Tropical TROPICAL PACIFIC GALLERY PACIFIC TROPICAL Main Entrance TROPICAL PACIFIC GALLERY PACIFIC TROPICAL Tropical Preview Tropical CHAPERONE …at the Aquarium Chaperones: Use this guide to move your group through the Gift Store Gift Store • Touch a shark Aquarium’s galleries. The background information, GUIDEguided questions, and activities will keep your • See a show students engaged and actively learning. Sea Otters Honda Theater Sea Otter Honda Theater Sea Otter • Visit a Discovery Lab • Ask questions Surge Channel Surge • Have fun! Kelp NORTHERN PACIFIC GALLERY PACIFIC NORTHERN NORTHERN PACIFIC GALLERY PACIFIC NORTHERN Amber Forest Pool Blue Cavern Blue Cavern Cafe Ray Scuba Blue Ray Pool Cavern Cafe Scuba Ray Pool SOUTHERN CALIFORNIA/BAJA GALLERY SOUTHERN CALIFORNIA/BAJA SOUTHERN CALIFORNIA/BAJA GALLERY SOUTHERN CALIFORNIA/BAJA Seals & Sea Lions Seals & Sea Lions Seals & Sea Lions …back at school Seals & Sea Lions • Write or draw about your trip to the Aquarium Ecosystems • Consider a classroom animal adoption Kelp vs. Coral • Visit aquariumofpacific.org/teachers Welcome to the Kelp Forest and Coral Reef! Shark Lagoon • Keep learning more These unique ecosystems each have producers, Forest like plants and kelp, which take energy from the Lorikeet Shark sun to make their own food, and consumers, Lagoon Shark Lagoon Forest which are incredible animals that rely on producers Lorikeet Blue Cavern, Amber Forest, Kelp Camouflage, Camouflage, Kelp Amber Forest, Blue Cavern, Channel, Sea Surge Otters Ray Pool, and other consumers for food. -

Kelp Forest Seasons of the Sea 89 ©2001 the Regents of the University of California
KELP FOREST SEASONS OF THE SEA FOR THE TEACHER Discipline Earth Science Themes Patterns of Change Key Concept Like land habitats, a kelp forest goes through seasonal changes that effect the animals and plants within the community. Synopsis Students work in groups to act out the seasonal changes and yearly variations that effect the life within a kelp forest. Science Process Skills communicating, comparing, organizing, relating, inferring Social Skills cooperating, sharing and attentive listening Vocabulary benthic, canopy, kelp wrack, herbivore, holdfast, photic zone, phytoplankton, stipe, upwelling, zooplankton MATERIALS Into the Activities Visuals depicting kelp forests: • videos- ("Forests of the Sea", "Ocean Realm", and NOVA'S "Kelp Forest" are some good ones) • slides/or pictures- (check out from MARE library or purchase from Monterey Bay Aquarium) Kelp Forest Seasons of the Sea 89 ©2001 The Regents of the University of California • books- (see literature list, check out from MARE library or purchase from the Monterey Bay Aquarium) butcher, poster or flip chart paper or sentence strips Through the Activities For creating costumes or props: • scissors, glue, staplers • colored construction, tissue or other paper • misc. materials-as needed Beyond the Activities • scenes #1-6 (remove numbers) from each act/copied onto index cards. 1 set for each group of 5-6 students • string or masking tape • butcher, poster or flip chart paper • colored markers or paint INTRODUCTION We often think of marine habitats as being unaffected by the seasonal changes that we experience on land. While seasonal variations in ocean temperature are not as extreme as those experienced on land (e.g., a hot summer day may not effect sea temperature at all), there are seasonal changes within marine ecosystems that have great impact on the plants and animals within them. -

Biodiversity of Kelp Forests and Coralline Algae Habitats in Southwestern Greenland
diversity Article Biodiversity of Kelp Forests and Coralline Algae Habitats in Southwestern Greenland Kathryn M. Schoenrock 1,2,* , Johanne Vad 3,4, Arley Muth 5, Danni M. Pearce 6, Brice R. Rea 7, J. Edward Schofield 7 and Nicholas A. Kamenos 1 1 School of Geographical and Earth Sciences, University of Glasgow, Gregory Building, Lilybank Gardens, Glasgow G12 8QQ, UK; [email protected] 2 Botany and Plant Science, National University of Ireland Galway, Ryan Institute, University Rd., H91 TK33 Galway, Ireland 3 School of Engineering, Geosciences, Infrastructure and Society, Heriot-Watt University, Riccarton Campus, Edinburgh EH14 4AS, UK; [email protected] 4 School of Geosciences, Grant Institute, University of Edinburgh, Edinburgh EH28 8, UK 5 Marine Science Institute, The University of Texas at Austin, College of Natural Sciences, 750 Channel View Drive, Port Aransas, TX 78373-5015, USA; [email protected] 6 Department of Biological and Environmental Sciences, School of Life and Medical Sciences, University of Hertfordshire, Hatfield, Hertfordshire AL10 9AB, UK; [email protected] 7 Geography & Environment, School of Geosciences, University of Aberdeen, Elphinstone Road, Aberdeen AB24 3UF, UK; [email protected] (B.R.R.); j.e.schofi[email protected] (J.E.S.) * Correspondence: [email protected]; Tel.: +353-87-637-2869 Received: 22 August 2018; Accepted: 22 October 2018; Published: 25 October 2018 Abstract: All marine communities in Greenland are experiencing rapid environmental change, and to understand the effects on those structured by seaweeds, baseline records are vital. The kelp and coralline algae habitats along Greenland’s coastlines are rarely studied, and we fill this knowledge gap for the area around Nuuk, west Greenland. -
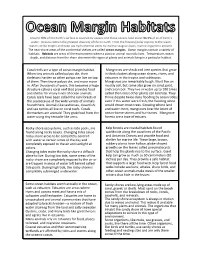
Mangroves Are Shrub and Tree Species That Grow in Thick Clusters Along
Around 70% of the Earth’s surface is covered by oceans and those oceans hold about 96.5% of all of Earth’s water. Oceans contain the greatest diversity of life on Earth. From the freezing polar regions to the warm waters of the tropics and deep sea hydrothermal vents to shallow seagrass beds, marine organisms abound. The near-shore areas of the continental shelves are called ocean margins. Ocean margins contain a variety of habitats. Habitats are areas of the environment where a plant or animal normally lives. Temperature, ocean depth, and distance from the shore determine the types of plants and animals living in a particular habitat. Coral reefs are a type of ocean margin habitat. Mangroves are shrub and tree species that grow When tiny animals called polyps die, their in thick clusters along ocean shores, rivers, and skeletons harden so other polyps can live on top estuaries in the tropics and subtropics. of them. Then those polyps die, and more move Mangroves are remarkably tough. Most live on in. After thousands of years, this becomes a huge muddy soil, but some also grow on sand, peat, structure called a coral reef that provides food and coral rock. They live in water up to 100 times and shelter for many kinds of ocean animals. saltier than most other plants can tolerate. They Corals reefs have been called the rain forests of thrive despite twice-daily flooding by ocean tides; the sea because of the wide variety of animals even if this water were fresh, the flooding alone found there. -

'Submerged Mangrove Forests Are Scientific Wonders — and Home To
TIMES EVOKE ‘Submerged mangrove forests are scientific wonders — and home to rich biodiversity’ n ocean. And a forest. It might seem that when two such forces of nature meet, one Graeme P Berlyn is EH Harriman Professor of Forest Management and Physiology of Trees at Yale University’s School of water roots. These vertical roots are Aoverwhelms the other. However, the op- called pneumatophores because they posite is true. When oceans and forests meet, Forestry and Environmental Studies. Writing for Times Evoke, the distinguished environmental expert discusses the complex contain pores (lenticels) on their sur- their worlds mingle and merge together in science of submerged mangroves, spread across coastlines from Africa to Asia — and home to thousands of species: face that can take up oxygen. They extraordinary harmony. The balance achieved Photos: Getty Images also produce less developed seedlings between these elements then creates a stunning angrove forests comprise on their branches. These are shed and panorama of life. Submerged forests feature the the interface between wet- can float for a time but these will not world’s greatest biodiversity, each layer blossom- lands and sea grass mead- complete seedling development un- ing with living beings, from bacteria and phyto- Mows along a vast expanse less they find muddy or solid ground. plankton, fish and worms, all sheltered within of tropical shorelines all over the Black mangrove roots can only filter wet woods, to shorebirds and migratory flocks world. They also occur along bays, es- out about 90% of the salt in its envi- nestled in branches and over-ground roots, to tuaries or mouths of rivers by these ronment, but it can expel the extra animals, from tiny tree-climbing crabs to mighty shores. -

Global Kelp Forest Restoration: Past Lessons, Status, and Future Goals
Title: Global Kelp Forest Restoration: Past lessons, status, and future goals Running Title: global kelp forest restoration List of Authors: Aaron Eger1*, Ezequiel M. Marzinelli2,3,4, Hartvig Christie5, Camilla W. Fagerli5, Daisuke Fujita6, Alejandra Paola Gonzalez7, Seokwoo Hong8, Jeong Ha Kim8, Lynn Chi Lee9,10, Tristin Anoush McHugh11,12, Gregory N. Nishihara13, Masayuki Tatsumi14, Peter D. Steinberg1, 3, Adriana Vergés1, 3 1 Centre for Marine Science and Innovation & Ecology and Evolution Research Centre, School of Biological, Earth and Environmental Sciences, The University of New South Wales, Sydney, NSW 2052, Australia 2 The University of Sydney, School of Life and Environmental Sciences, Sydney, NSW 2006, Australia 3 Sydney Institute of Marine Science, 19 Chowder Bay Rd, Mosman, NSW 2088, Australia 4 Singapore Centre for Environmental Life Sciences Engineering, Nanyang Technological University, Singapore 637551, Singapore 5 Norwegian Institute for Water Research, Gaustadallèen 21, 0349 Oslo, Norway 6 University of Tokyo Marine Science and Technology, School of Marine Bioresources, Applied Phycology, Konan, Minato-ku, Tokyo, Japan 7 Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile. Las Palmeras 3425, Ñuñoa, Santiago. Chile 8 Department of Biological Sciences, Sungkyunkwan University, Suwon, South Korea 9 Gwaii Haanas National Park Reserve, National Marine Conservation Area Reserve, and Haida Heritage Site, 60 Second Beach Road, Skidegate, Haida Gwaii, British Columbia, V0T 1S1, Canada 10 Canada & School -

Management and Protection of Kelp Forest Systems
Olympic Coast National Marine Sanctuary Management and Protection of Kelp Forest Systems Management Issue Kelp habitat and the associated ecological community need to be monitored to assess long-term trends for management needs and resource protection in the Olympic Coast National Marine Sanctuary (OCNMS or Sanctuary). Description Giant kelp (Macrocystis integrifola) and bull kelp (Nereocystis leutkeana) are the dominant kelp species in the nearshore waters of the Sanctuary. Kelps can grow up to 20 meters in length, and form dense patches known as kelp forests which provide habitat for a wide variety of organisms. Kelp forests are often compared to tree forests on land since both have distinct layers: a sunny canopy at the surface, a dimly lit middle, and a dim forest bottom. This pattern of vertical zonation provides a variety of microhabitats or niches that support a large and Kelp forest as fish habitat. Photo credit: Steve Fisher diverse assemblage of biological species. Sea otters often forage and rest in kelp canopies, while many fish and invertebrate species find safe cover within the intricate structure of the kelp forest. The complex structure of this living habitat buffers nearshore areas from the force of waves, surges and currents, thereby creating a relatively protected environment. The kelp canopy is dynamic and has considerate inter- annual variability, yet these kelp forests are generally thought to be stable. The overall trend is increased total surface area of floating kelp along the outer coast and the Straits of Juan -

Marine Ecology Progress Series 400:283
Vol. 400: 283–288, 2010 MARINE ECOLOGY PROGRESS SERIES Published February 11 doi: 10.3354/meps08425 Mar Ecol Prog Ser NOTE Purple sea urchins Strongylocentrotus purpuratus reduce grazing rates in response to risk cues from the spiny lobster Panulirus interruptus Catherine M. Matassa* Marine Science Center, Northeastern University, 430 Nahant Road, Nahant, Massachusetts 01908, USA ABSTRACT: The classical view of trophic cascades is that predators, by consuming herbivores, exert a positive indirect effect on plants. Although this form of trophic cascade has been demonstrated in a variety of terrestrial, aquatic, and marine systems, growing evidence suggests many trophic cascades are driven by anti-predator behaviors in prey. Despite abundant evidence of behavioral responses by sea urchins to predators, there has been little examination of how predation risk may influence urchin grazing rates. To determine if purple sea urchins Strongylocentrotus purpuratus graze less in the presence of predation risk, I monitored individual urchin grazing on the kelp Macrocystis pyrifera in the presence and absence of waterborne cues from damaged conspecifics and the predatory spiny lobster Panulirus interruptus. Sea urchin grazing rates were similar in the presence and absence of damaged conspecifics, but sea urchins exposed to lobster cues, regardless of lobster diet, reduced grazing rates by 44%. Given that trophic cascades involving herbivorous sea urchins exert an impor- tant influence on primary production in kelp forests, these results suggest that predation risk may play an important but under-appreciated role in the dynamics of kelp forest food webs and primary production. KEY WORDS: Non-consumptive effect · Trait-mediated indirect interactions · Trophic cascade · Kelp forest · Macrocystis pyrifera Resale or republication not permitted without written consent of the publisher INTRODUCTION et al.