Diet Comparison of Coexisting Collared Owlet (Glaucidium Brodiei
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

S.No Institutename Enrolled Appeared Passed Promoted Fail Absent Percen
FEDERAL DIRECTORATE OF EDUCATION EXAMINATION SECTION Centralized Annual Examination Class-V,2017 **** INSTITUTION WISE RESULT BRIEFPASS/PROMOTED/FAILED/PASS PERCENTAGE/GRADES/GPA S.No InstituteName Enrolled Appeared Passed Promoted Fail Absent Percen. GPA 1 Islamabad College for Boys G-6/3, Islamabad. 349 348 314 32 2 1 99.43 4.18 2 Islamabad Model College for Boys F-7/3, Islamabad. 184 180 162 15 3 4 98.33 3.68 3 Islamabad Model College for Boys F-8/4, Islamabad. 172 171 158 12 1 1 99.42 4.50 4 Islamabad Model College for Boys I-8/3, Islamabad. 129 128 106 18 4 1 96.88 3.41 5 Islamabad Model College for Boys I-10/1, Islamabad. 145 145 142 3 0 0 100.00 4.54 6 Islamabad Model College for Boys G-10/4, Islamabad. 196 194 192 2 0 2 100.00 4.82 7 Islamabad Model College for Boys G-11/1, Islamabad. 75 75 54 15 6 0 92.00 2.79 8 Islamabad Model School ( I-V ) G-11/1, Islamabad. 65 65 50 15 0 0 100.00 2.85 9 Islamabad Model College for Boys F-10/3, Islamabad. 74 73 65 6 2 1 97.26 3.82 10 Islamabad Model School ( I-V ) F-10/1, Islamabad. 26 26 21 4 1 0 96.15 3.65 11 Islamabad Model School ( I-V ) F-10/2, Islamabad. 37 37 31 4 2 0 94.59 3.16 12 Islamabad Model School ( I-V ) F-10/4, Islamabad. -

Roosting Ecology of Spotted Owlet (Athene Brama) and Barn Owl (Tyto
Journal of Entomology and Zoology Studies 2018; 6(1): 829-833 E-ISSN: 2320-7078 P-ISSN: 2349-6800 Roosting ecology of spotted owlet (Athene brama) JEZS 2018; 6(1): 829-833 © 2018 JEZS and barn owl (Tyto alba) in agroecosystem of Received: 04-11-2017 Accepted: 05-12-2017 Punjab Yashika Gaba Department of Zoology, Punjab Agricultural University, Yashika Gaba and Nisha Vashishat Ludhiana, Punjab, India Abstract Nisha Vashishat The Spotted Owlet (Athene brama) and Barn Owl (Tyto alba) are the most common owl species in Department of Zoology, Punjab Punjab. The roosting ecology of Spotted Owlet and Barn Owl was observed at different sites of Punjab Agricultural University, Ludhiana, Punjab, India Agricultural University, villages Baranhara and Ladhowal of district Ludhiana. The preferred habitat of owls in selected locations was observed to record their utilization for roosting. Different trees utilized for roosting by Spotted Owlet were mulberry (Morus spp.), mango (Mangifera indica), banyan (Ficus spp.), peepal (Ficus religiosa) and dek (Melia azedarach). Barn Owl preferred peepal and jamun trees for roosting. Barn Owl also preferred closed grain sheds and empty building crevices for roosting. Mean tree height was observed to be 13.00±1.22 m at Punjab Agricultural University, 15.33±2.85 m at Baranhara and 12.50±0.50 m at Ladhowal. Most preferred tree utilized for roosting by Spotted Owlet was mulberry and dek tree and that of Barn Owl was peepal tree. Seasonal variation was also observed in roosting behaviour. Spotted Owlets were seen shifting from one roosting sites to another or spending more time in their cavities during the winter season. -

Estimations Relative to Birds of Prey in Captivity in the United States of America
ESTIMATIONS RELATIVE TO BIRDS OF PREY IN CAPTIVITY IN THE UNITED STATES OF AMERICA by Roger Thacker Department of Animal Laboratories The Ohio State University Columbus, Ohio 43210 Introduction. Counts relating to birds of prey in captivity have been accomplished in some European countries; how- ever, to the knowledge of this author no such information is available in the United States of America. The following paper consistsof data related to this subject collected during 1969-1970 from surveys carried out in many different direc- tions within this country. Methods. In an attempt to obtain as clear a picture as pos- sible, counts were divided into specific areas: Research, Zoo- logical, Falconry, and Pet Holders. It became obvious as the project advanced that in some casesthere was overlap from one area to another; an example of this being a falconer working with a bird both for falconry and research purposes. In some instances such as this, the author has used his own judgment in placing birds in specific categories; in other in- stances received information has been used for this purpose. It has also become clear during this project that a count of "pets" is very difficult to obtain. Lack of interest, non-coop- eration, or no available information from animal sales firms makes the task very difficult, as unfortunately, to obtain a clear dispersal picture it is from such sourcesthat informa- tion must be gleaned. However, data related to the importa- tion of birds' of prey as recorded by the Bureau of Sport Fisheries and Wildlife is included, and it is felt some observa- tions can be made from these figures. -

Biodiversity Chapter Three
CHAPTER THREE BIODIVERSITY CHAPTER THREE BIODIVERSITY 3.1 The term ‘biodiversity’ encompasses preservation of biodiversity is both a matter the variety of life on Earth. It is defined as of investment and insurance to a) sustain the variability among living organisms and and improve agricultural, forestry and the ecological complexes of which they are fisheries production, b) act as a buffer part, including diversity within and between against harmful environmental changes, c) species and ecosystems. Biodiversity provide raw materials for scientific and manifests at species genetic and ecosystem industrial innovations, and d) safe guard levels. Biodiversity has direct consumptive transferring biological richness to future value in food, agriculture, medicine, generations. industry, etc. It also has aesthetic and recreational value. 3.5 Biodiversity the world over is in peril because the habitats are threatened due 3.2 India is one of the 12 mega- to such development programmes as biodiversity countries of the world. From creation of reservoirs, mining, forest about 70% of the total geographical area clearing, laying of transport and surveyed so far, 46,000 plant species and communication networks, etc. It is 81,000 animal species representing about estimated that in the world wide 7% of the world’s flora and 6.5% of the perspective, slightly over 1000 animal world's fauna, respectively, have been species and sub-species are threatened with described. Out of the total twelve an extinction rate of one per year, while biodiversity hot spots in the world, India has 20,000 flowering plants are thought to be at two; one is the north east region and other risk. -

Biodiversity Profile of Afghanistan
NEPA Biodiversity Profile of Afghanistan An Output of the National Capacity Needs Self-Assessment for Global Environment Management (NCSA) for Afghanistan June 2008 United Nations Environment Programme Post-Conflict and Disaster Management Branch First published in Kabul in 2008 by the United Nations Environment Programme. Copyright © 2008, United Nations Environment Programme. This publication may be reproduced in whole or in part and in any form for educational or non-profit purposes without special permission from the copyright holder, provided acknowledgement of the source is made. UNEP would appreciate receiving a copy of any publication that uses this publication as a source. No use of this publication may be made for resale or for any other commercial purpose whatsoever without prior permission in writing from the United Nations Environment Programme. United Nations Environment Programme Darulaman Kabul, Afghanistan Tel: +93 (0)799 382 571 E-mail: [email protected] Web: http://www.unep.org DISCLAIMER The contents of this volume do not necessarily reflect the views of UNEP, or contributory organizations. The designations employed and the presentations do not imply the expressions of any opinion whatsoever on the part of UNEP or contributory organizations concerning the legal status of any country, territory, city or area or its authority, or concerning the delimitation of its frontiers or boundaries. Unless otherwise credited, all the photos in this publication have been taken by the UNEP staff. Design and Layout: Rachel Dolores -
![Explorer Research Article [Tripathi Et Al., 6(3): March, 2015:4304-4316] CODEN (USA): IJPLCP ISSN: 0976-7126 INTERNATIONAL JOURNAL of PHARMACY & LIFE SCIENCES (Int](https://docslib.b-cdn.net/cover/4638/explorer-research-article-tripathi-et-al-6-3-march-2015-4304-4316-coden-usa-ijplcp-issn-0976-7126-international-journal-of-pharmacy-life-sciences-int-1074638.webp)
Explorer Research Article [Tripathi Et Al., 6(3): March, 2015:4304-4316] CODEN (USA): IJPLCP ISSN: 0976-7126 INTERNATIONAL JOURNAL of PHARMACY & LIFE SCIENCES (Int
Explorer Research Article [Tripathi et al., 6(3): March, 2015:4304-4316] CODEN (USA): IJPLCP ISSN: 0976-7126 INTERNATIONAL JOURNAL OF PHARMACY & LIFE SCIENCES (Int. J. of Pharm. Life Sci.) Study on Bird Diversity of Chuhiya Forest, District Rewa, Madhya Pradesh, India Praneeta Tripathi1*, Amit Tiwari2, Shivesh Pratap Singh1 and Shirish Agnihotri3 1, Department of Zoology, Govt. P.G. College, Satna, (MP) - India 2, Department of Zoology, Govt. T.R.S. College, Rewa, (MP) - India 3, Research Officer, Fishermen Welfare and Fisheries Development Department, Bhopal, (MP) - India Abstract One hundred and twenty two species of birds belonging to 19 orders, 53 families and 101 genera were recorded at Chuhiya Forest, Rewa, Madhya Pradesh, India from all the three seasons. Out of these as per IUCN red list status 1 species is Critically Endangered, 3 each are Vulnerable and Near Threatened and rest are under Least concern category. Bird species, Gyps bengalensis, which is comes under Falconiformes order and Accipitridae family are critically endangered. The study area provide diverse habitat in the form of dense forest and agricultural land. Rose- ringed Parakeets, Alexandrine Parakeets, Common Babblers, Common Myna, Jungle Myna, Baya Weavers, House Sparrows, Paddyfield Pipit, White-throated Munia, White-bellied Drongo, House crows, Philippine Crows, Paddyfield Warbler etc. were prominent bird species of the study area, which are adapted to diversified habitat of Chuhiya Forest. Human impacts such as Installation of industrial units, cutting of trees, use of insecticides in agricultural practices are major threats to bird communities. Key-Words: Bird, Chuhiya Forest, IUCN, Endangered Introduction Birds (class-Aves) are feathered, winged, two-legged, Birds are ideal bio-indicators and useful models for warm-blooded, egg-laying vertebrates. -

Strigiformes) and Lesser Nighthawks (Chodeiles Acutipennis
UNIVERSITY OF CALIFORNIA RIVERSIDE The Evolution of Quiet Flight in Owls (Strigiformes) and Lesser Nighthawks (Chodeiles acutipennis) A Dissertation submitted in partial satisfaction of the requirements for the degree of Doctor of Philosophy in Evolution, Ecology, and Organismal Biology by Krista Le Piane December 2020 Dissertation Committee: Dr. Christopher J. Clark, Chairperson Dr. Erin Wilson Rankin Dr. Khaleel A. Razak Copyright by Krista Le Piane 2020 The Dissertation of Krista Le Piane is approved: Committee Chairperson University of California, Riverside ACKNOWLEDGEMENTS I thank my Oral Exam Committee: Dr. Khaleel A. Razak (chairperson), Dr. Erin Wilson Rankin, Dr. Mark Springer, Dr. Jesse Barber, and Dr. Scott Curie. Thank you to my Dissertation Committee: Dr. Christopher J. Clark (chairperson), Dr. Erin Wilson Rankin, and Dr. Khaleel A. Razak for their encouragement and help with this dissertation. Thank you to my lab mates, past and present: Dr. Sean Wilcox, Dr. Katie Johnson, Ayala Berger, David Rankin, Dr. Nadje Najar, Elisa Henderson, Dr. Brian Meyers Dr. Jenny Hazelhurst, Emily Mistick, Lori Liu, and Lilly Hollingsworth for their friendship and support. I thank the Natural History Museum of Los Angeles County (LACM), the California Academy of Sciences (CAS), Museum of Vertebrate Zoology (MVZ) at UC Berkeley, the American Museum of Natural History (ANMH), and the Natural History Museum (NHM) in Tring for access to specimens used in Chapter 1. I would especially like to thank Kimball Garrett and Allison Shultz for help at LACM. I also thank Ben Williams, Richard Jackson, and Reddit user NorthernJoey for permission to use their photos in Chapter 1. Jessica Tingle contributed R code and advice to Chapter 1 and I would like to thank her for her help. -

List of Ngos
Registration Sr. Name of NGO Address Status(Reg No,& No Date & Relevant Law Malot Commercial Center, Shahpur VSWA/ICT/494 ,Date: 1. Aas Welfare Association Simly Dam Road, Islamabad. 051- 6/9/2006, VSWA 2232394, 03005167142 Ordinance 1961 VSWA/ICT/439, Village Khana Dak, Madina Town, Date:31/3/2004, 2. Abaseen Welfare Society Jabbah, Barma Chowk, Islamabad. VSWA Ordinance 03005308511 1961 Office No.5, 1st Floor, A & K Plaza, F- VSWA/ICT/511,Date 3. Acid Survivors Foundation. 10 Markaz, Islamabad. 051-2214052, 02/08/2007, VSWA 03008438984 Ordinance 1961 VSWA/ICT/282, Date Ali Pur Frash Lehtrar Road, 4. Akhmat Welfare Centre 24/04/1998, VSWA Islamabad. 051-2519062 Ordinance 1961 Al Mustafa Towers VSWA/ICT/438, Date # 204, Al Mustafa Towers F-10 5. Residents Associations 27/03/2004,VSWA Markaz, Islamabad. 03458555872 (AMPTRWA) Ordinance 1961 VSWA/ICT/520, Date Al-Firdous Welfare Mohallah Ara, Bharakau, Islamabad. 6. 2/04/2008, VSWA Association 051-2512308, 03008565227 Ordinance 1961 VSWA/ICT/261,Date:0 Model Village Ali Pur Farash 7. Al-Itehad Welfare Society 5/08/1996, VSWA Islamabad. Ordinance 1961 VSWA/ICT/202,Date:1 NearF.G.Boys High School Shahdra 8. Al-Jihad Youth Organization 7/06/1993,VSWA Islamabad Ordinance 1961 VSWA/ICT/377, Date H.No.39 Gali No.67 I- 9. Al-Noor Foundation. 17.4.2002, VSWA 10/1,Islamabad.051-4443227 Ordinance 1961 VSWA/ICT/90, Date 10. Al-Noor Islamic Center Rawal Town Islamabad 2.12.1990 VSWA Ordinance 1961 VSWA/ICT/89, Date 11. Al-Noor Islamic Center Sharifabad Tarlai Islamabad 2.12.1990 VSWA Ordinance 1961 VSWA/ICT/271, Date PEC Building, Ataturk Avenue, G-5/2, 12. -
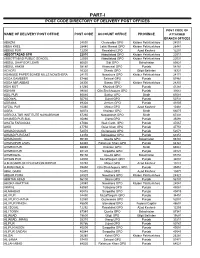
Part-I: Post Code Directory of Delivery Post Offices
PART-I POST CODE DIRECTORY OF DELIVERY POST OFFICES POST CODE OF NAME OF DELIVERY POST OFFICE POST CODE ACCOUNT OFFICE PROVINCE ATTACHED BRANCH OFFICES ABAZAI 24550 Charsadda GPO Khyber Pakhtunkhwa 24551 ABBA KHEL 28440 Lakki Marwat GPO Khyber Pakhtunkhwa 28441 ABBAS PUR 12200 Rawalakot GPO Azad Kashmir 12201 ABBOTTABAD GPO 22010 Abbottabad GPO Khyber Pakhtunkhwa 22011 ABBOTTABAD PUBLIC SCHOOL 22030 Abbottabad GPO Khyber Pakhtunkhwa 22031 ABDUL GHAFOOR LEHRI 80820 Sibi GPO Balochistan 80821 ABDUL HAKIM 58180 Khanewal GPO Punjab 58181 ACHORI 16320 Skardu GPO Gilgit Baltistan 16321 ADAMJEE PAPER BOARD MILLS NOWSHERA 24170 Nowshera GPO Khyber Pakhtunkhwa 24171 ADDA GAMBEER 57460 Sahiwal GPO Punjab 57461 ADDA MIR ABBAS 28300 Bannu GPO Khyber Pakhtunkhwa 28301 ADHI KOT 41260 Khushab GPO Punjab 41261 ADHIAN 39060 Qila Sheikhupura GPO Punjab 39061 ADIL PUR 65080 Sukkur GPO Sindh 65081 ADOWAL 50730 Gujrat GPO Punjab 50731 ADRANA 49304 Jhelum GPO Punjab 49305 AFZAL PUR 10360 Mirpur GPO Azad Kashmir 10361 AGRA 66074 Khairpur GPO Sindh 66075 AGRICULTUR INSTITUTE NAWABSHAH 67230 Nawabshah GPO Sindh 67231 AHAMED PUR SIAL 35090 Jhang GPO Punjab 35091 AHATA FAROOQIA 47066 Wah Cantt. GPO Punjab 47067 AHDI 47750 Gujar Khan GPO Punjab 47751 AHMAD NAGAR 52070 Gujranwala GPO Punjab 52071 AHMAD PUR EAST 63350 Bahawalpur GPO Punjab 63351 AHMADOON 96100 Quetta GPO Balochistan 96101 AHMADPUR LAMA 64380 Rahimyar Khan GPO Punjab 64381 AHMED PUR 66040 Khairpur GPO Sindh 66041 AHMED PUR 40120 Sargodha GPO Punjab 40121 AHMEDWAL 95150 Quetta GPO Balochistan 95151 -

Bird Observations from Valmiki Tiger Reserve, Bihar Anwaruddin Choudhury
CHOUDHURY: Valmiki Tiger Reserve 57 Bird observations from Valmiki Tiger Reserve, Bihar Anwaruddin Choudhury Choudhury, A. U., 2016. Bird observations from Valmiki Tiger Reserve, Bihar. Indian BIRDS 11 (3): 57–63. Anwaruddin Choudhury, Secretary to the Government of Assam, House No. 7, Islampur Road, Near Gate No. 1 of Nehru Stadium, Guwahati 781007, Assam, India. E-mail: [email protected] Manuscript received on 23 November 2015. Abstract Valmiki Tiger Reserve, in Bihar, is a relatively poorly known bird area. Observations from a recent visit (16 October to 06 November 2015) are documented here. Noteworthy records include some globally threatened, and near-threatened species such as Lesser Adjutant Leptoptilos javanica, Swamp Francolin Francolinus gularis, Oriental Darter Anhinga melanogaster, and River Lapwing Vanellus duvauceli. There were records of White-backed Gyps bengalensis and Slender-billed Vultures G. tenuirostris, and Sarus Crane Grus antigone. Historical records of the Bengal Florican Houbaropsis bengalensis, from the general area, also exist. A large roost of migratory Amur Falcons Falco amurensis was also discovered just outside the reserve. Several species could have been reported as new for Valmiki Tiger Reserve, but lack of published materials has made it difficult to identify these. Altogether 246 species of birds were listed in this work although there could be more than 300 species. Most of the summer visitors, such as cuckoos, and winter visitors, such as ducks, and waders, could not be observed owing to the time of visit. Details of some important observations are presented. Conservation issues are discussed briefly, and recommendations are made for the protection of habitats, and birds. -

Ghana Included in the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES)
IDENTIFICATION GUIDE The Species of Ghana Included in the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) YEAR 2018 IDENTIFICATION GUIDE The CITES Species of Ghana Born Free USA thanks the National Oceanic and Atmospheric Administration (NOAA) for funding this guide and the Ghana authorities for their support. See the last section for a list of useful contacts, including the organizations displayed above. PHOTOS: MICHAEL HEYNS, BROCKEN INAGLORY, GEORGE CHERNILEVSKY, ALEX CHERNIKH, HANS HILLEWAERT, DAVID D’O, JAKOB FAHR TABLE OF CONTENTS How to use this guide ..........................................1 CHORDATA / ELASMOBRANCHII What is CITES? ..............................................3 / Carcharhiniformes ........................................101 What is the IUCN Red List? .....................................10 / Lamniformes .............................................101 How to read this guide ........................................13 / Orectolobiformes .........................................102 What the IUCN colors mean ....................................15 / Pristiformes ..............................................103 Steps for CITES permits .......................................17 Presentation of shark and ray species listed in CITES in West Africa ........19 CHORDATA / ACTINOPTERI / Syngnathiformes ..........................................103 CHORDATA / MAMMALIA / Artiodactyla ..............................................51 ARTHROPODA / ARACHNIDA / Carnivora ................................................53 -

The Rediscovery of the Forest Owlet Athene (Heteroglaux) Blewitti
FORKTAIL 14 (1998): 53-55 The rediscovery of the Forest Owlet Athene (Heteroglaux) blewitti BEN F. KING AND PAMELA C. RASMUSSEN One hundred and thirteen years after the last genuine record, the Forest Owlet Athene blewitti has been rediscovered in low foothills dry-deciduous forest north of Shahada, Maharashtra, India. Two individuals were observed in the same locality from 25–27 November 1997, exhibiting strong diurnality and behaving in a relatively conspicuous, confiding manner. Their identification was verified by extensive videotape and photographs, and these field observations confirm that the Forest Owlet is a readily identified species. The Forest Owlet Athene (Heteroglaux) blewitti is known from just seven specimens, collected from four sites in central India from 1872 to 1884. For the next 113 years there were no genuine records of the species. Several unsuccessful searches for the bird were made in recent years, including by Sálim Ali, S. Dillon Ripley and colleagues. A number of ornithologists considered the species possibly extinct (see Rasmussen and Collar 1998 for historical review). From 13–27 November 1997 we (along with D. F. Abbott) searched for the Forest Owlet in central India in the states of southeastern Madhya Pradesh, westernmost Orissa and northwestern Maharashtra, in forests as near as possible to the sites where it had been collected over a century ago. However, the original collection sites could be located only approximately. See Appendix for details of areas visited. On 25 November we found a single Forest Owlet in Habitat at the rediscovery site at the beginning of the rainy north-western Maharashtra, at 460 m elevation, north season.