Marine Ecology Progress Series 370:45
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Algae & Marine Plants of Point Reyes
Algae & Marine Plants of Point Reyes Green Algae or Chlorophyta Genus/Species Common Name Acrosiphonia coalita Green rope, Tangled weed Blidingia minima Blidingia minima var. vexata Dwarf sea hair Bryopsis corticulans Cladophora columbiana Green tuft alga Codium fragile subsp. californicum Sea staghorn Codium setchellii Smooth spongy cushion, Green spongy cushion Trentepohlia aurea Ulva californica Ulva fenestrata Sea lettuce Ulva intestinalis Sea hair, Sea lettuce, Gutweed, Grass kelp Ulva linza Ulva taeniata Urospora sp. Brown Algae or Ochrophyta Genus/Species Common Name Alaria marginata Ribbon kelp, Winged kelp Analipus japonicus Fir branch seaweed, Sea fir Coilodesme californica Dactylosiphon bullosus Desmarestia herbacea Desmarestia latifrons Egregia menziesii Feather boa Fucus distichus Bladderwrack, Rockweed Haplogloia andersonii Anderson's gooey brown Laminaria setchellii Southern stiff-stiped kelp Laminaria sinclairii Leathesia marina Sea cauliflower Melanosiphon intestinalis Twisted sea tubes Nereocystis luetkeana Bull kelp, Bullwhip kelp, Bladder wrack, Edible kelp, Ribbon kelp Pelvetiopsis limitata Petalonia fascia False kelp Petrospongium rugosum Phaeostrophion irregulare Sand-scoured false kelp Pterygophora californica Woody-stemmed kelp, Stalked kelp, Walking kelp Ralfsia sp. Silvetia compressa Rockweed Stephanocystis osmundacea Page 1 of 4 Red Algae or Rhodophyta Genus/Species Common Name Ahnfeltia fastigiata Bushy Ahnfelt's seaweed Ahnfeltiopsis linearis Anisocladella pacifica Bangia sp. Bossiella dichotoma Bossiella -

And Red Sea Urchins
NEGATIVELY CORRELATED ABUNDANCE SUGGESTS COMPETITION BETWEEN RED ABALONE (Haliotis rufescens) AND RED SEA URCHINS (Mesocentrotus franciscanus) INSIDE AND OUTSIDE ESTABLISHED MPAs CLOSED TO COMMERCIAL SEA URCHIN HARVEST IN NORTHERN CALIFORNIA By Johnathan Centoni A Thesis Presented to The Faculty of Humboldt State University In Partial Fulfillment of the Requirements for the Degree Master of Science in Biology Committee Membership Dr. Sean Craig, Committee Chair Dr. Brian Tissot, Committee Member Dr. Paul Bourdeau, Committee Member Dr. Joe Tyburczy, Committee Member Dr. Erik Jules, Program Graduate Coordinator May 2018 ABSTRACT NEGATIVELY CORRELATED ABUNDANCE SUGGESTS COMPETITION BETWEEN RED ABALONE (Haliotis rufescens) AND RED SEA URCHINS (Mesocentrotus franciscanus) INSIDE AND OUTSIDE ESTABLISHED MPAs CLOSED TO COMMERCIAL SEA URCHIN HARVEST IN NORTHERN CALIFORNIA Johnathan Centoni Red abalone and sea urchins are both important herbivores that potentially compete with each other for resources like food and space along the California coast. Red abalone supported a socioeconomically important recreational fishery during this study (which was closed in 2018) and red sea urchins support an important commercial fishery. Both red sea urchins and red abalone feed on the same macroalgae (including Pterygophora californica, Laminaria setchellii, Stephanocystis osmundacea, Costaria costata, Alaria marginata, Nereocystis leutkeana), and a low abundance of this food source during the period of this project may have created a highly competitive environment for urchins and abalone. Evidence that suggests competition between red abalone and red sea urchins can be seen within data collected during the years of this study (2014-2016): a significantly higher red sea urchin density, concomitant with a significantly lower red abalone density, was observed within areas closed to commercial sea urchin harvest (in MPAs) compared to nearby reference areas open to sea urchin harvest. -

Biomechanical Analysis of Wave-Induced Mortality in the Marine Alga Pterygophora Californica
MARINE ECOLOGY - PROGRESS SERIES Vol. 36: 163-170, 1987 Published March 2 Mar. Ecol. hog. Ser. Biomechanical analysis of wave-induced mortality in the marine alga Pterygophora californica Richard F. Biedka, John M. Gosline & Robert E. De Wreede Departments of Zoology and Botany, University of British Columbia, Vancouver, British Columbia V6T 2B1, Canada ABSTRACT: A biomechanical analysis was carried out on the stipe of the marine brown alga Pterygophora cahfornica (Ruprecht). Tensile and compressive mechanical tests on the cortical material revealed that the plant is constructed from a reasonably stiff (tensile modulus of elasticity = 60 MN m-2), strong (tensile strength = 9 MN m-2) and extensible (fdure strain = 0.49) material. Consequently, the stipe is a very robust structure that is extremely difficult to break. However, the work of fracture of the cortical material is only about 400 J m-', making it a relatively brittle material. Thus, it is expected that small notches or flaws at the stipe surface will dramatically reduce the ability of the stipe to resist bending forces caused by waves. Fracture mechanics analysis suggests a critical flaw size for the cortical material of about 0.2 mm, and any surface defect larger than this size will degrade the mechanical strength of the plant. The red urchin Strongylocentrotus franciscanus is known to graze on the stipe of P. californica. This grazing is known to mtroduce surface flaws an order of magnitude or more larger than the critical flaw size, and therefore it IS very likely that such grazlng will make a substantial contribution to wave-induced mortahty of these plants. -

Patterns of Septal Biomineralization in Scleractinia Compared with Their 28S Rrna Phylogeny
PBlackwell Publishingatterns Ltd. of septal biomineralization in Scleractinia compared with their 28S rRNA phylogeny: a dual approach for a new taxonomic framework JEAN-PIERRE CUIF, GUILLAUME LECOINTRE, CHRISTINE PERRIN, ANNIE TILLIER & SIMON TILLIER Accepted: 2 December 2002 Cuif, J.-P., Lecointre, G., Perrin, C., Tillier, A. & Tillier, S. (2003). Patterns of septal bio- mineralization in Scleractinia compared with their 28S rRNA phylogeny: a dual approach for a new taxonomic framework. — Zoologica Scripta, 32, 459–473. A molecular phylogeny of the Scleractinia is reconstructed from approximately 700 nucleo- tides of the 5′end of the 28S rDNA obtained from 40 species. A comparison of molecular phylogenic trees with biomineralization patterns of coral septa suggests that at least five clades are corroborated by both types of data. Agaricidae and Dendrophylliidae are found to be monophyletic, that is supported by microstructural data. Conversely, Faviidae and Caryophyl- liidae are found to be paraphyletic: Cladocora should be excluded from the faviids, whereas Eusmilia should be excluded from the caryophylliids. The conclusion is also supported by the positions, sizes and shapes of centres of calcification. The traditional Guyniidae are diphyletic, corroborating Stolarski’s hypothesis ‘A’. Some results from our most parsimonious trees are not strongly statistically supported but corroborated by other molecular studies and micro- structural observations. For example, in the scleractinian phylogenetic tree, there are several lines of evidence (including those from our data) to distinguish a Faviidae–Mussidae lineage and a Dendrophylliidae–Agaricidae–Poritidae–Siderastreidae lineage. From a methodological standpoint, our results suggest that co-ordinated studies creating links between biomineralization patterns and molecular phylogeny may provide an efficient working approach for a re- examination of scleractinian classification. -

Abundance and Clonal Replication in the Tropical Corallimorpharian Rhodactis Rhodostoma
Invertebrate Biology 119(4): 351-360. 0 2000 American Microscopical Society, Inc. Abundance and clonal replication in the tropical corallimorpharian Rhodactis rhodostoma Nanette E. Chadwick-Furmana and Michael Spiegel Interuniversity Institute for Marine Science, PO. Box 469, Eilat, Israel, and Faculty of Life Sciences, Bar Ilan University, Ramat Gan, Israel Abstract. The corallimorpharian Rhodactis rhodostoma appears to be an opportunistic species capable of rapidly monopolizing patches of unoccupied shallow substrate on tropical reefs. On a fringing coral reef at Eilat, Israel, northern Red Sea, we examined patterns of abundance and clonal replication in R. rhodostoma in order to understand the modes and rates of spread of polyps across the reef flat. Polyps were abundant on the inner reef flat (maximum 1510 polyps m-* and 69% cover), rare on the outer reef flat, and completely absent on the outer reef slope at >3 m depth. Individuals cloned throughout the year via 3 distinct modes: longitudinal fission, inverse budding, and marginal budding. Marginal budding is a replicative mode not previously described. Cloning mode varied significantly with polyp size. Approximately 9% of polyps cloned each month, leading to a clonal doubling time of about 1 year. The rate of cloning varied seasonally and depended on day length and seawater temperature, except for a brief reduction in cloning during midsummer when polyps spawned gametes. Polyps of R. rhodo- stoma appear to have replicated extensively following a catastrophic low-tide disturbance in 1970, and have become an alternate dominant to stony corals on parts of the reef flat. Additional key words: Cnidaria, coral reef, sea anemone, asexual reproduction, Red Sea Soft-bodied benthic cnidarians such as sea anemo- & Chadwick-Furman 1999a). -

Factors Affecting the Abundance of Paracyathus Stearns!! on Subtidal Rock Walls
MLML / M8~RllIBR;jRY 8272 MOSS LANDING RD. MOSS LANDING, CA 95039 FACTORS AFFECTING THE ABUNDANCE OF PARACYATHUS STEARNS!! ON SUBTIDAL ROCK WALLS by Mark Pranger A thesis submitted in partial fulfillment ofthe requirements for the degree of Master ofScience in Marine Science in the School ofNatural Sciences California State University, Fresno August 1999 ACKNOWLEDGMENTS I would like to thank the members of my graduate advisory committee for their advice and guidance during this process. Dr. James Nybakken for his interest and education in invertebrate biology and for allowing me to help in the teaching of others. For the insights and humor of Dr. Mike Foster, that helped improve this study and my education. For the help of Dr. Stephen Ervin who in the last hours help me through all CSU Fresno's paper work and deadlines. I am grateful to the staff at the Pollution Studies Lab at Granite Canyon for their help during experiments and for allowing me the use of their working space. I am indebted to the staff at Moss Landing Marine Labs that kept all the equipment working and operational, and to the Dr. Earl and Ethel Myers Oceanographic and Marine Biology Trust for helping to fund this project. Most of all, thanks goes to the many students at Moss Landing. Without their help this study would not have been completed. Special thanks goes to; Mat Edwards for being a faithful dive buddy, to Torno Eguchi, Michelle White, Bryn Phillips, Cassandra Roberts, Michelle Lander, and Lara Lovera for their help on statistics and editing of early drafts, and especially to Michele Jacobi for her help in all aspects of this study and her constant encouragement for me to finish. -

The Diet and Predator-Prey Relationships of the Sea Star Pycnopodia Helianthoides (Brandt) from a Central California Kelp Forest
THE DIET AND PREDATOR-PREY RELATIONSHIPS OF THE SEA STAR PYCNOPODIA HELIANTHOIDES (BRANDT) FROM A CENTRAL CALIFORNIA KELP FOREST A Thesis Presented to The Faculty of Moss Landing Marine Laboratories San Jose State University In Partial Fulfillment of the Requirements for the Degree Master of Arts by Timothy John Herrlinger December 1983 TABLE OF CONTENTS Acknowledgments iv Abstract vi List of Tables viii List of Figures ix INTRODUCTION 1 MATERIALS AND METHODS Site Description 4 Diet 5 Prey Densities and Defensive Responses 8 Prey-Size Selection 9 Prey Handling Times 9 Prey Adhesion 9 Tethering of Calliostoma ligatum 10 Microhabitat Distribution of Prey 12 OBSERVATIONS AND RESULTS Diet 14 Prey Densities 16 Prey Defensive Responses 17 Prey-Size Selection 18 Prey Handling Times 18 Prey Adhesion 19 Tethering of Calliostoma ligatum 19 Microhabitat Distribution of Prey 20 DISCUSSION Diet 21 Prey Densities 24 Prey Defensive Responses 25 Prey-Size Selection 27 Prey Handling Times 27 Prey Adhesion 28 Tethering of Calliostoma ligatum and Prey Refugia 29 Microhabitat Distribution of Prey 32 Chemoreception vs. a Chemotactile Response 36 Foraging Strategy 38 LITERATURE CITED 41 TABLES 48 FIGURES 56 iii ACKNOWLEDGMENTS My span at Moss Landing Marine Laboratories has been a wonderful experience. So many people have contributed in one way or another to the outcome. My diving buddies perse- vered through a lot and I cherish our camaraderie: Todd Anderson, Joel Thompson, Allan Fukuyama, Val Breda, John Heine, Mike Denega, Bruce Welden, Becky Herrlinger, Al Solonsky, Ellen Faurot, Gilbert Van Dykhuizen, Ralph Larson, Guy Hoelzer, Mickey Singer, and Jerry Kashiwada. Kevin Lohman and Richard Reaves spent many hours repairing com puter programs for me. -

Kelp Forest Monitoring Handbook — Volume 1: Sampling Protocol
KELP FOREST MONITORING HANDBOOK VOLUME 1: SAMPLING PROTOCOL CHANNEL ISLANDS NATIONAL PARK KELP FOREST MONITORING HANDBOOK VOLUME 1: SAMPLING PROTOCOL Channel Islands National Park Gary E. Davis David J. Kushner Jennifer M. Mondragon Jeff E. Mondragon Derek Lerma Daniel V. Richards National Park Service Channel Islands National Park 1901 Spinnaker Drive Ventura, California 93001 November 1997 TABLE OF CONTENTS INTRODUCTION .....................................................................................................1 MONITORING DESIGN CONSIDERATIONS ......................................................... Species Selection ...........................................................................................2 Site Selection .................................................................................................3 Sampling Technique Selection .......................................................................3 SAMPLING METHOD PROTOCOL......................................................................... General Information .......................................................................................8 1 m Quadrats ..................................................................................................9 5 m Quadrats ..................................................................................................11 Band Transects ...............................................................................................13 Random Point Contacts ..................................................................................15 -

The Earliest Diverging Extant Scleractinian Corals Recovered by Mitochondrial Genomes Isabela G
www.nature.com/scientificreports OPEN The earliest diverging extant scleractinian corals recovered by mitochondrial genomes Isabela G. L. Seiblitz1,2*, Kátia C. C. Capel2, Jarosław Stolarski3, Zheng Bin Randolph Quek4, Danwei Huang4,5 & Marcelo V. Kitahara1,2 Evolutionary reconstructions of scleractinian corals have a discrepant proportion of zooxanthellate reef-building species in relation to their azooxanthellate deep-sea counterparts. In particular, the earliest diverging “Basal” lineage remains poorly studied compared to “Robust” and “Complex” corals. The lack of data from corals other than reef-building species impairs a broader understanding of scleractinian evolution. Here, based on complete mitogenomes, the early onset of azooxanthellate corals is explored focusing on one of the most morphologically distinct families, Micrabaciidae. Sequenced on both Illumina and Sanger platforms, mitogenomes of four micrabaciids range from 19,048 to 19,542 bp and have gene content and order similar to the majority of scleractinians. Phylogenies containing all mitochondrial genes confrm the monophyly of Micrabaciidae as a sister group to the rest of Scleractinia. This topology not only corroborates the hypothesis of a solitary and azooxanthellate ancestor for the order, but also agrees with the unique skeletal microstructure previously found in the family. Moreover, the early-diverging position of micrabaciids followed by gardineriids reinforces the previously observed macromorphological similarities between micrabaciids and Corallimorpharia as -

Ibdiocc- Scor Wg
PROPOSAL FOR IBDIOCC- SCOR WG Submitted to: Dr. Edward Urban, Executive Secretary, Scientific Committee for Oceanic Research (SCOR) Submitted by: Dr. Robert Y. George, President, George Institute for Biodiversity and Sustainability (GIBS), 1320 Vanagrif Ct., Wake Forest, North Carolina. Date of Submission: April 15, 2016. IBDIOCC Interaction Between Drivers Impacting Ocean Carbonate Chemistry: How can Deep-Sea Coral Ecosystems respond to ASH/CSH Shoaling in Seamounts that pose imminent threats from Ocean Acidification? Summary/Abstract: We propose a new SCOR Working Group IBDIOCC (2017 to 2019) that seeks to assess new impacts on seamount ecosystems from ocean acidification (OA), that essentially looks at the impact of shoaling of ASH and CSH on the biota that include communities/species associated with deep sea scleractinian corals e.g. Lophelia pertusa and Solenosmilia variabilis) The WG, with members from both southern and northern hemispheres, seeks to re-evaluate and augment the science priorities defined in 2012 by the Census of the Marine Life, but taking into account the new climate change threats and challenges from shifts in ocean carbonate chemistry. The WG will incorporate recommendations from ‘Ocean In High Carbon World-Ocean Acidification international symposium which will be participated by Dr. George (chairman of WG) who will also present a paper on vulnerable deep sea ecosystems to ocean carbonate chemistry, especially seamounts southeast of Australia and New Zealand. The WG plans to develop a follow-on capacity building workshop in the ASLO annual meeting in Hawaii (2017) and in the AGU Ocean Sciences meeting in Portland, Oregon (2018). In 2017, the WG will meet for three days in 2017 at the ASLO annual meeting to generate two open-access publications; 1) the first global assessment of OA on seamount fauna, and 2) a peer-reviewed multi-authored paper to be submitted to NATURE CLIMATE. -
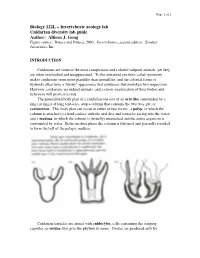
Cnidarians Are Some of the Most Conspicuous and Colorful Tidepool Animals, Yet They Are Often Overlooked and Unappreciated
Page 1 of 1 Biology 122L – Invertebrate zoology lab Cnidarian diversity lab guide Author: Allison J. Gong Figure source: Brusca and Brusca, 2003. Invertebrates, second edition. Sinauer Associates, Inc. INTRODUCTION Cnidarians are some of the most conspicuous and colorful tidepool animals, yet they are often overlooked and unappreciated. To the untrained eye their radial symmetry makes cnidarians seem more plantlike than animallike, and the colonial forms of hydroids often have a "bushy" appearance that reinforces that mistaken first impression. However, cnidarians are indeed animals, and a closer examination of their bodies and behaviors will prove it to you. The generalized body plan of a cnidarian consists of an oral disc surrounded by a ring (or rings) of long tentacles, atop a column that contains the two-way gut, or coelenteron. This body plan can occur in either of two forms: a polyp, in which the column is attached to a hard surface with the oral disc and tentacles facing into the water; and a medusa, in which the column is (usually) unattached and the entire organism is surrounded by water. In the medusa phase the column is flattened and generally rounded to form the bell of the pelagic medusa. Cnidarian tentacles are armed with cnidocytes, cells containing the stinging capsules, or cnidae, that give the phylum its name. Cnidae are produced only by Page 2 of 2 cnidarians, although they can occasionally be found in the cerata of nudibranch molluscs that feed on cnidarians: through a process that is not understood, some nudibranchs are able to ingest cnidae from their prey and sequester them, unfired, in their cerata for defense against their own predators. -

CNIDARIA Corals, Medusae, Hydroids, Myxozoans
FOUR Phylum CNIDARIA corals, medusae, hydroids, myxozoans STEPHEN D. CAIRNS, LISA-ANN GERSHWIN, FRED J. BROOK, PHILIP PUGH, ELLIOT W. Dawson, OscaR OcaÑA V., WILLEM VERvooRT, GARY WILLIAMS, JEANETTE E. Watson, DENNIS M. OPREsko, PETER SCHUCHERT, P. MICHAEL HINE, DENNIS P. GORDON, HAMISH J. CAMPBELL, ANTHONY J. WRIGHT, JUAN A. SÁNCHEZ, DAPHNE G. FAUTIN his ancient phylum of mostly marine organisms is best known for its contribution to geomorphological features, forming thousands of square Tkilometres of coral reefs in warm tropical waters. Their fossil remains contribute to some limestones. Cnidarians are also significant components of the plankton, where large medusae – popularly called jellyfish – and colonial forms like Portuguese man-of-war and stringy siphonophores prey on other organisms including small fish. Some of these species are justly feared by humans for their stings, which in some cases can be fatal. Certainly, most New Zealanders will have encountered cnidarians when rambling along beaches and fossicking in rock pools where sea anemones and diminutive bushy hydroids abound. In New Zealand’s fiords and in deeper water on seamounts, black corals and branching gorgonians can form veritable trees five metres high or more. In contrast, inland inhabitants of continental landmasses who have never, or rarely, seen an ocean or visited a seashore can hardly be impressed with the Cnidaria as a phylum – freshwater cnidarians are relatively few, restricted to tiny hydras, the branching hydroid Cordylophora, and rare medusae. Worldwide, there are about 10,000 described species, with perhaps half as many again undescribed. All cnidarians have nettle cells known as nematocysts (or cnidae – from the Greek, knide, a nettle), extraordinarily complex structures that are effectively invaginated coiled tubes within a cell.