Download This PDF File
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Summary of Offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019
Summary of offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019 3841 Number of items in BX 301 thru BX 463 1815 Number of unique text strings used as taxa 990 Taxa offered as bulbs 1056 Taxa offered as seeds 308 Number of genera This does not include the SXs. Top 20 Most Oft Listed: BULBS Times listed SEEDS Times listed Oxalis obtusa 53 Zephyranthes primulina 20 Oxalis flava 36 Rhodophiala bifida 14 Oxalis hirta 25 Habranthus tubispathus 13 Oxalis bowiei 22 Moraea villosa 13 Ferraria crispa 20 Veltheimia bracteata 13 Oxalis sp. 20 Clivia miniata 12 Oxalis purpurea 18 Zephyranthes drummondii 12 Lachenalia mutabilis 17 Zephyranthes reginae 11 Moraea sp. 17 Amaryllis belladonna 10 Amaryllis belladonna 14 Calochortus venustus 10 Oxalis luteola 14 Zephyranthes fosteri 10 Albuca sp. 13 Calochortus luteus 9 Moraea villosa 13 Crinum bulbispermum 9 Oxalis caprina 13 Habranthus robustus 9 Oxalis imbricata 12 Haemanthus albiflos 9 Oxalis namaquana 12 Nerine bowdenii 9 Oxalis engleriana 11 Cyclamen graecum 8 Oxalis melanosticta 'Ken Aslet'11 Fritillaria affinis 8 Moraea ciliata 10 Habranthus brachyandrus 8 Oxalis commutata 10 Zephyranthes 'Pink Beauty' 8 Summary of offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019 Most taxa specify to species level. 34 taxa were listed as Genus sp. for bulbs 23 taxa were listed as Genus sp. for seeds 141 taxa were listed with quoted 'Variety' Top 20 Most often listed Genera BULBS SEEDS Genus N items BXs Genus N items BXs Oxalis 450 64 Zephyranthes 202 35 Lachenalia 125 47 Calochortus 94 15 Moraea 99 31 Moraea -

GTAC/CBPEP/ EU Project on Employment-Intensive Rural Land Reform in South Africa: Policies, Programmes and Capacities
GTAC/CBPEP/ EU project on employment-intensive rural land reform in South Africa: policies, programmes and capacities Municipal case study Matzikama Local Municipality, Western Cape David Mayson, Rick de Satgé and Ivor Manuel with Bruno Losch Phuhlisani NPC March 2020 Abbreviations and acronyms BEE Black Economic Empowerment CASP Comprehensive Agricultural Support Programme CAWH Community Animal Health Worker CEO Chief Executive Officer CPA Communal Property of Association CPAC Commodity Project Allocation Committee DAAC District Agri-Park Advisory Committee DAPOTT District Agri Park Operational Task Team DoA Department of Agriculture DRDLR Department of Rural Development and Land Reform DWS Department of Water and Sanitation ECPA Ebenhaeser CPA FALA Financial Assistance Land FAO Food and Agriculture Organisation FPSU Farmer Production Support Unit FTE Full-Time Equivalent GGP Gross Geographic Product GDP Gross Domestic Product GVA Gross Value Added HDI Historically Disadvantaged Individual IDP Integrated Development Plan ILO International Labour Organisation LED Local economic development LORWUA Lower Olifants Water Users Association LSU Large stock units NDP National Development Plan PDOA Provincial Department of Agriculture PGWC Provincial Government of the Western Cape PLAS Proactive Land Acquisition Strategy SDF Spatial Development Framework SLAG Settlement and Land Acquisition Grant SSU Small stock unit SPP Surplus People Project TRANCRAA Transformation of Certain Rural Areas Act WUA Water Users Association ii Table of Contents -

Partial Flora Survey Rottnest Island Golf Course
PARTIAL FLORA SURVEY ROTTNEST ISLAND GOLF COURSE Prepared by Marion Timms Commencing 1 st Fairway travelling to 2 nd – 11 th left hand side Family Botanical Name Common Name Mimosaceae Acacia rostellifera Summer scented wattle Dasypogonaceae Acanthocarpus preissii Prickle lily Apocynaceae Alyxia Buxifolia Dysentry bush Casuarinacea Casuarina obesa Swamp sheoak Cupressaceae Callitris preissii Rottnest Is. Pine Chenopodiaceae Halosarcia indica supsp. Bidens Chenopodiaceae Sarcocornia blackiana Samphire Chenopodiaceae Threlkeldia diffusa Coast bonefruit Chenopodiaceae Sarcocornia quinqueflora Beaded samphire Chenopodiaceae Suada australis Seablite Chenopodiaceae Atriplex isatidea Coast saltbush Poaceae Sporabolis virginicus Marine couch Myrtaceae Melaleuca lanceolata Rottnest Is. Teatree Pittosporaceae Pittosporum phylliraeoides Weeping pittosporum Poaceae Stipa flavescens Tussock grass 2nd – 11 th Fairway Family Botanical Name Common Name Chenopodiaceae Sarcocornia quinqueflora Beaded samphire Chenopodiaceae Atriplex isatidea Coast saltbush Cyperaceae Gahnia trifida Coast sword sedge Pittosporaceae Pittosporum phyliraeoides Weeping pittosporum Myrtaceae Melaleuca lanceolata Rottnest Is. Teatree Chenopodiaceae Sarcocornia blackiana Samphire Central drainage wetland commencing at Vietnam sign Family Botanical Name Common Name Chenopodiaceae Halosarcia halecnomoides Chenopodiaceae Sarcocornia quinqueflora Beaded samphire Chenopodiaceae Sarcocornia blackiana Samphire Poaceae Sporobolis virginicus Cyperaceae Gahnia Trifida Coast sword sedge -

Radiation in the Cape Flora and the Phylogeny of Peacock Irises Moraea
ARTICLE IN PRESS MOLECULAR PHYLOGENETICS AND EVOLUTION Molecular Phylogenetics and Evolution xxx (2002) xxx–xxx www.academicpress.com Radiation in the Cape flora and the phylogeny of peacock irises Moraea (Iridaceae) based on four plastid DNA regions Peter Goldblatt,a Vincent Savolainen,b,* Obie Porteous,b Ivan Sostaric,b Martyn Powell,b Gail Reeves,c John C. Manning,c Timothy G. Barraclough,d and Mark W. Chaseb a Missouri Botanical Garden, P.O. Box 299, St. Louis, Missouri 63166, USA b Molecular Systematics Section, Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond TW9 3DS, UK c National Botanical Institute, Kirstenbosch, Private Bag X7, Cape Town, South Africa d Department of Biology and NERC Centre for Population Biology, Imperial College, Silwood Park, Ascot, Berkshire SL5 7PY, UK Received 2 February 2002; received in revised form 22 April 2002 Abstract Phylogenetic analyses of four plastid DNA regions, the rbcL exon, trnL intron, trnL–trnF intergenic spacer, and rps16 intron from each of 73 species in the African genus Moraea (Iridaceae: Irideae) including accessions of all major species clusters in the genus, show Moraea to be paraphyletic when Barnardiella, Galaxia, Hexaglottis, Homeria (all southern African), and Gynandriris (Eurasian as well) were recognized as separate genera. There are several small, isolated species clusters at the basal nodes of the tree that are all restricted to the winter-rainfall zone of southern Africa (the Greater Cape floral kingdom) and a few, highly derived, large species groups that have radiated extensively within the winter-rainfall zone. Mapping of floral traits shows that an Iris-type flower is ancestral in Moraea. -

15. Contribuciones29
Acta BotanicaBotanica Malacitana Malacitana 29. 29: 2004 297-315 Málaga, 2004 297 CONTRIBUCIONES A LA FLORA VASCULAR DE ANDALUCÍA (ESPAÑA) (96-100) 96. A NEW SPECIES OF IRIDACEAE FOR SW OF SPAIN Enrique SÁNCHEZ GULLÓN & Pedro WEICKERT Una nueva especie de Iridaceae para el sudoeste de España. Palabras clave. Flora, corología, Ferraria crispa, Iridaceae, xenófito adventicio naturalizado, Huelva, Andalucía Key words. Flora, corology, Ferraria crispa, Iridaceae, xenophyte adventitious naturalised, Huelva, Andalusia A study of the flora and vegetation of is cited only one species, Ferraria crispa province of Huelva was carried out, mainly in Burman. Nova Acta Acad. Caesar. Leopold.- the Paraje Natural Marismas del Odiel and the Carol. 2:199 (1761)= Ferraria undulata L. Sp. litoral (Sánchez Gullón et al., 1997; Sánchez ed. 2 p. 1353 (1763). This specimen was Gullón et al., 1998; Sánchez Gullón, 1999; introduced in Europe in 1640, where is cited Cobos et al., 2003; etc.). In this note it includes always adventitious near of garden coast in observation of vascular plant with chorological Portugal and east of Spain [Lu.: CW and SW. and ecological references of a new alien species Hs.:Mll] (Hutchinson, J., 1946; Walter et al., for this province. 1986; Amaral Franco & Rocha Afonso, 1994; The Ferraria Burman [Ferraria L. Bólos et al., 1990). Also is cited in N Africa (1753); Ferraria Miller] is a genera the (Maire, 1971), and the Macaronesica Region Iridaceae family of aprox. 10 especies natural in Canary Island (Hierro) and Madeira of South Africa and Tropical (cf.Vos, M. P. The (Hansen, A. & Sunding, P., 1985). Our research African genum Ferraria, Journal of South note expands the to-date known distribution up African Botany 45: 295-376 (1979). -

City of Mandurah Environmental Weeds Black Flag Ferraria Crispa (Burm.) FAMILY: IRIDACEAE
City of Mandurah Environmental Weeds Black Flag Ferraria crispa (Burm.) FAMILY: IRIDACEAE SYNONYMS: FERRARIA UNDULATA ORIGIN: CAPE PROVINCE, SOUTH AFRICA OTHER NAMES: SPINEKOPBLOM, FLAG LILY GROWTH FORM: PERENNIAL CORM Photographer Roger Cousens Copyright © Western Australian Agriculture Authority, 1995 HABITAT: COASTAL WOODLAND, PLANT DESCRIPTION Black flag is a small perennial herb, with thick, succulent leaves and GRASSLANDS, HERBFIELDS, SAND flowering stems up to 450mm tall. Black Flag occurs in coastal heath, DUNES AND DISTURBED AREAS IN Tuart, Agonis and Banksia woodland from Perth to Cape Riche. It is often found growing in clumps and readily recognised by its succulent SANDY OR LOAM SOILS. foliage even when not in flower. Plants produce dark brown to black, foul-smelling Iris-type flowers from August to November. Flowers only DISPERSAL: last one day but are produced in succession. SOIL MOVEMENT, SURFACE IMPACTS RUNOFF Black Flag reproduces asexually from cormlets and is a prolific seeder, which can form monocultures under favorable conditions. Dense growth results in the smothering of smaller native herbs and prevents native FLOWERING PERIOD: plant recruitment. AUGUST– NOVEMBER SUGGESTED CONTROL METHOD Hand remove very small populations in degraded sites. Sift soil to find FIRE RESPONSE: all corms. In degraded sites try glyphosate 1% + metsulfuron methyl 0.2 UNDERGROUND CORMS SURVIVE g/15 L + Pulse®. Takes a number of years to control populations. Read the manufacturers’ labels and material safety data sheets before using FIRE -

Insights from Microsporogenesis in Asparagales
EVOLUTION & DEVELOPMENT 9:5, 460–471 (2007) Constraints and selection: insights from microsporogenesis in Asparagales Laurent Penet,a,1,Ã Michel Laurin,b Pierre-Henri Gouyon,a,c and Sophie Nadota aLaboratoire Ecologie, Syste´matique et Evolution, Batiment 360, Universite´ Paris-Sud, 91405 Orsay Ce´dex, France bUMR CNRS 7179, Universite´ Paris 6FPierre & Marie Curie, 2 place Jussieu, Case 7077, 75005 Paris, France cMuse´um National d’Histoire Naturelle, De´partement de Syste´matique et Evolution Botanique, 12 rue Buffon, 75005 Paris CP 39, France ÃAuthor for correspondence (email: [email protected]) 1Current address: Department of Biological Sciences, University of Pittsburgh, 4249 Fifth & Ruskin, Pittsburgh, PA 15260, USA. SUMMARY Developmental constraints have been proposed different characteristics of microsporogenesis, only cell to interfere with natural selection in limiting the available wall formation appeared as constrained. We show that set of potential adaptations. Whereas this concept has constraints may also result from biases in the correlated long been debated on theoretical grounds, it has been occurrence of developmental steps (e.g., lack of successive investigated empirically only in a few studies. In this article, cytokinesis when wall formation is centripetal). We document we evaluate the importance of developmental constraints such biases and their potential outcomes, notably the during microsporogenesis (male meiosis in plants), with an establishment of intermediate stages, which allow emphasis on phylogenetic patterns in Asparagales. Different development to bypass such constraints. These insights are developmental constraints were tested by character discussed with regard to potential selection on pollen reshuffling or by simulated distributions. Among the morphology. INTRODUCTION 1991) also hindered tests using the concept (Pigliucci and Kaplan 2000). -

43574 31-7 Legala.Pdf
Government Gazette Staatskoerant REPUBLIC OF SOUTH AFRICA REPUBLIEK VAN SUID-AFRIKA July Vol. 661 Pretoria, 31 2020 Julie No. 43574 PART 1 OF 2 LEGAL NOTICES A WETLIKE KENNISGEWINGS ISSN 1682-5843 N.B. The Government Printing Works will 43574 not be held responsible for the quality of “Hard Copies” or “Electronic Files” submitted for publication purposes 9 771682 584003 AIDS HELPLINE: 0800-0123-22 Prevention is the cure 2 No. 43574 GOVERNMENT GAZETTE, 31 JULY 2020 IMPORTANT NOTICE OF OFFICE RELOCATION Private Bag X85, PRETORIA, 0001 149 Bosman Street, PRETORIA Tel: 012 748 6197, Website: www.gpwonline.co.za URGENT NOTICE TO OUR VALUED CUSTOMERS: PUBLICATIONS OFFICE’S RELOCATION HAS BEEN TEMPORARILY SUSPENDED. Please be advised that the GPW Publications office will no longer move to 88 Visagie Street as indicated in the previous notices. The move has been suspended due to the fact that the new building in 88 Visagie Street is not ready for occupation yet. We will later on issue another notice informing you of the new date of relocation. We are doing everything possible to ensure that our service to you is not disrupted. As things stand, we will continue providing you with our normal service from the current location at 196 Paul Kruger Street, Masada building. Customers who seek further information and or have any questions or concerns are free to contact us through telephone 012 748 6066 or email Ms Maureen Toka at [email protected] or cell phone at 082 859 4910. Please note that you will still be able to download gazettes free of charge from our website www.gpwonline.co.za. -
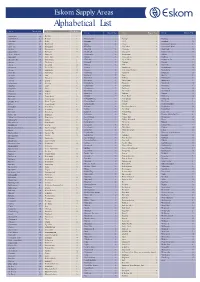
Alphabetical List
Eskom Supply Areas Alphabetical List Area Block No. Area Block No. Area Block No. Area Block No. Area Block No. De Rust 3 Abbotsdale 12 Admiral's Park 5 De Zalze 8 Klaarstroom 2 Nyanga 1 Starking 5 Adriaanse 13 Delft 1 Klapmuts 16 OCC 10 Steinkopf 10 AECI site 5 Dias Strand 7 Klawer 15 Okiep 10 Stellenbosch farmers 8 Agter Paarl 10 Doringbaai 15 Klein Brak 7 Olyfenhout 16 Stellenbosch Munic 2 Agulhas 10 Doringwater 10 Klein Pella 16 Omdraai 16 Steyrkraal 16 Airport City 14 Du Noon 14 Kleinbegin 16 Ongegund 2 Stikland farmers 15 Airport Industria 14 Duinzicht 7 Kleinmond 2 Onrusrivier 5 Stilbaai 7 Albertinia 7 Duiwelsnek 16 Klipdale 10 Ontseepkans 16 Stofkraal 15 Alexander Bay 10 Dysselsdorp 3 Klipheuwel 10 Op die Berg 13 Stompneus Bay 13 Algeria 15 Ebenhaeser 15 Kliprand 15 Osplaas 4 Storage 5 Alheit 16 Eendekuil 15 Kliprug 8 OTB 10 Stormsvlei 10 Alkantrant 10 Eersterivier 2 Knysna 7 Oudtshoorn 3 Strandfontein 15 Amcor 13 Elandsbaai 15 Koegas 16 Outeniqua Farmers 3 Strausburg 16 Arabella 3 Elandskloof 13 Koekenaap 15 Paardevlei 5 Struisbaai 10 Arniston 10 Elim 3 Koekoeb 16 Paarl 9 Sun City 5 Ashton 3 Elsiesriver 13 Koelenhof 8 Padkloof 16 Sunningdale 14 Askraal 10 Esethu 5 Komaggas 10 Paleisheuwel 15 Sutherland 4 Atlantis rural 10 Esterhof 11 Koringberg 3 Panorama 2 Suurbraak 10 Atlas Park 14 Faure 8 Kotzehoop 10 Papendorp 15 Swartberg 4 Augrabies 16 Fibre 16 Krantshoek 7 Parklands 14 Swartdraai 16 Aurora 15 Firgrove 8 Krantzkop 11 Paternoster 6 Swellendam 10 Avonwood 13 Firlands 5 Kuilsrivier 6 Paulputs 16 Taaiput 16 Barandas 3 Fisantekraal -

A Review Ofthe Namaqua Gecko, Pachydactylus Namaquensis (Reptilia: Gekkonidae) from Southern Africa, with the Description of Two New Species
S. Afr. J. Zool. 1996,31(2) 53--69 A review ofthe Namaqua gecko, Pachydactylus namaquensis (Reptilia: Gekkonidae) from southern Africa, with the description of two new species William R. Branch' Department of Herpetology, Port Elizabeth Museum, P.O. Box 13147, Humewood, 6013 South Africa Aaron M, Bauer Biology Department, Villanova University, 800 Lancaster Avenue, Villanova, Pennsylvania 19085, USA David A. Good Museum of Natural Science, 119 Foster Hall, Louisiana State University, Baton Rouge, Louisiana 70803, USA Received 19 September 1995; accepted 22 January 1996 An analysis of morphological and allozyme variation in the Namaqua gecko, Pachydactylus namaquensis from southern Africa is presented. Three separately evolving lineages, well defined by morphology and allozyme var iation, are identified. The isolated southern population, occurring on the southern escarpment and Cape Fold Mountains surrounding the western Litlle Karoo, is named P. kladaroderrna sp. nov., and is characterized by a slit-like ear opening, low number of granules bordering the mental (3--6) and mental and adjacent infralabials (5- 13), the frequent (79%) occurence of the supralabial entering the nostril, and its drab brown base colouration. A northern population, occurring in southern Namibia and the Richtersveld is named P. haackei sp. nov" and is characterized by its more rounded or squared ear opening; high number of granules bordering the mental and adjacent infralabials (11-19), the general exclusion of the supralabial from the nostril (only 3.7% entry), and brighter, lighter colouration. It is further differentiated from P. kladaroderrna on the basis of fixed differences at 11 allozyme loci. Both new species differ from P. -

Threatened Ecosystems in South Africa: Descriptions and Maps
Threatened Ecosystems in South Africa: Descriptions and Maps DRAFT May 2009 South African National Biodiversity Institute Department of Environmental Affairs and Tourism Contents List of tables .............................................................................................................................. vii List of figures............................................................................................................................. vii 1 Introduction .......................................................................................................................... 8 2 Criteria for identifying threatened ecosystems............................................................... 10 3 Summary of listed ecosystems ........................................................................................ 12 4 Descriptions and individual maps of threatened ecosystems ...................................... 14 4.1 Explanation of descriptions ........................................................................................................ 14 4.2 Listed threatened ecosystems ................................................................................................... 16 4.2.1 Critically Endangered (CR) ................................................................................................................ 16 1. Atlantis Sand Fynbos (FFd 4) .......................................................................................................................... 16 2. Blesbokspruit Highveld Grassland -

Groundwater Overview for Lower Orange Water Management Area
ISP: LOWER ORANGE WMA Appendix B Groundwater Overview for Lower Orange Water Management Area 03 - APPENDIX B GROUNDWATER.DOC JULY 2004 B1 DEVELOPMENT OF INTERNAL STRATEGIC PERSPECTIVES GROUNDWATER OVERVIEW FOR LOWER ORANGE CATCHMENT MANAGEMENT AREA Prepared by: JA Pretorius and I Dennis Darcy Groundwater Scientists and Consultants Bloemfontein Prepared for: Directorate Water Resource Planning DWAF Private Bag X313 Pretoria SEPTEMBER 2003 TABLE OF CONTENTS 1 OVERARCHING ISSUES ........................................................................................................ 3 1.1 AVAILABILITY OF GROUNDWATER INFORMATION IN THE CATCHMENT AREA............................... 3 1.2 OVERVIEW OF GROUNDWATER RESOURCES AND USE THROUGHOUT THE CATCHMENT AREA .... 3 1.2.1 Industrial and mining.................................................................................................. 3 1.2.2 Agriculture.................................................................................................................. 3 1.2.3 Domestic .................................................................................................................... 3 1.3 GROUNDWATER QUALITY IN THE CATCHMENT AREA ............................................................... 4 1.3.1 Natural ....................................................................................................................... 4 1.3.2 Point and diffusive pollution ....................................................................................... 7 1.4 GROUNDWATER MANAGEMENT