Equus Zebra Zebra) T
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Conservation Goals for the Cape Mountain Zebra Equus Zebra Zebra—Security in Numbers?
Conservation goals for the Cape mountain zebra Equus zebra zebra—security in numbers? H ALSZKA H RABAR and G RAHAM I. H. KERLEY Abstract The target of the 2002 IUCN Action Plan for the required to achieve the conservation objectives. However, Cape mountain zebra Equus zebra zebra was for a popu- such compromised targets may also be influenced by a poor lation of 2,500 animals. We assessed the validity of this goal understanding of the needs of a species for survival, as well by reviewing the conservation status of the subspecies and as underestimation of the potential to achieve larger, more the prospects of achieving larger populations. All subpopu- reliable, conservation targets. An example of the former is lations were identified and data on each were collected by where conservation targets for many large mammal species means of a questionnaire survey. The total extant Cape have been set based on earlier estimates of minimum viable mountain zebra population was found to consist of at least populations with an effective population size of c. 500 (e.g. 2,790 individuals in 52 subpopulations. The target number Lande & Barrowclough, 1987; Armbruster & Lande, 1993). of 2,500 has therefore been exceeded and this success is This figure of 500 is, however, based on maintaining genetic largely attributable to the private sector, as there are at variation in leg hair patterns in fruit flies and is 30 years old present double the number of privately owned subpopu- (Franklin, 1980) and outdated. Recent estimates have been lations (35) compared to formally protected subpopulations consistently higher than this (Reed et al., 2003). -

Ecology and Habitat Suitability of Cape Mountain Zebra (Equus Zebra Zebra) in the Western Cape, South Africa
Ecology and habitat suitability of Cape mountain zebra (Equus zebra zebra) in the Western Cape, South Africa by Adriaan Jacobus Olivier Thesis presented in partial fulfilment of the requirements for the degree Masters of Science at Stellenbosch University Department of Conservation Ecology and Entomology, Faculty of AgriScience Supervisor: Dr Alison J. Leslie Co-supervisor: Dr Jason I. Ransom December 2019 Stellenbosch University https://scholar.sun.ac.za Declaration By submitting this thesis electronically, I declare that the entirety of the work contained therein is my own, original work, that I am the sole author thereof (save to the extent explicitly otherwise stated), that reproduction and publication thereof by Stellenbosch University will not infringe any third party rights and that I have not previously in its entirety or in part submitted it for obtaining any qualification. Jaco Olivier December 2019 Copyright © 2019 Stellenbosch University All rights reserved ii Stellenbosch University https://scholar.sun.ac.za Abstract Endemic to South Africa, the Cape mountain zebra (Equus zebra zebra) historically occurred throughout the Western Cape, and parts of the Northern and Eastern Cape. However, due to human impacts fewer than 50 individuals remained by the 1950’s. Conservation efforts over the past 50 years have resulted in the population increasing to over 4700 individuals and having moved on the IUCN red list, from Critically Endangered to Least Concern. As there are still many isolated meta-populations, CapeNature established a Biodiversity Management Plan for the conservation of Cape mountain zebra in the Western Cape. In 2001, 15 (six males and nine females) Cape mountain zebra was reintroduced into Bakkrans Nature Reserve, situated in the Cederberg Wilderness Area of South Africa. -

Landman Etal. Habitat Suitability Assessements for Namaqualand
A habitat suitability assessment for the introduction of Black Rhinoceros and Mountain Zebra to the Namaqua National Park and Upland priority region in the Succulent Karoo Biome M. Landman, A.M. Shrader and G.I.H. Kerley Terrestrial Ecology Research Unit Nelson Mandela Metropolitan University Port Elizabeth Report No. C110 6031 May 2006 A habitat suitability assessment for the introduction of Black Rhinoceros and Mountain Zebra to the Namaqua National Park and Upland priority region in the Succulent Karoo Biome M. Landman, A.M. Shrader and G.I.H. Kerley Terrestrial Ecology Research Unit PO Box 77000 Nelson Mandela Metropolitan University Port Elizabeth, 6031 Report No. C110 May 2006 Report to: Conservation International Foundation - South Africa Kirstenbosch National Botanical Gardens Cape Town, 7735 CONTENTS Page 1. EXECUTIVE SUMMARY 1 2. TERMS OF REFERENCE 2 3. INTRODUCTION 3 4. PROPOSED AREAS 4 5. APPROACH 5 6. MOUNTAIN ZEBRA Equus zebra 6.1 Historical occurrence 9 6.2 Ecological requirements and general biology 10 6.3 Habitat suitability assessment 6.3.1 Platbakkies 11 6.3.2 Spoegriver 13 6.3.3 Rooiberg catchment 14 6.3.4 Namaqua National Park 17 6.4 Summary & Recommendations 20 6.4.1 Reintroduction strategy 21 6.4.2 Potential carrying capacity 22 7. BLACK RHINOCEROS Diceros bicornis 7.1 Historical occurrence 24 2.2 Ecological requirements and general biology 25 7.3 Habitat suitability assessment 7.3.1 Namaqua National Park 26 7.4 Summary & Recommendations 28 8. POTENTIAL FOR OTHER HERBIVORE RE-INTRODUCTIONS 28 9. ACKNOWLEDGEMENTS 29 10. REFERENCES 29 1. EXECUTIVE SUMMARY The aim of the study was to determine the potential suitability of areas in Namaqualand for the introduction of mountain zebra and black rhinoceros. -

Mountain Zebra Project – 8Th Progress Report Population Ecology of Hartmann’S Mountain Zebra
Mountain Zebra Project – 8th progress report Population ecology of Hartmann’s mountain zebra 2018 Waiting to drink at the Moringa waterhole in NamibRand Nature Reserve. The animals are members of a breeding group and the small waterhole is occupied by a dominant female (right). The two animals waiting to drink are offspring in the group, the youngest born in the 2016-17 summer and, on the far left, a male born in 2015. Photo © L. M. Gosling 1 Introduction This is the eighth progress report on a long-term study of the population ecology and conservation of Hartmann’s mountain zebra (Equus zebra hartmannae), through wet and dry cycles, that has been underway since 2005 in partnership with the Ministry of Environment and Tourism, Gondwana Canyon Park and NamibRand Nature Reserve. It deals mainly with 2017, the most recent year of almost complete sampling. An additional notable event was the completion of the IUCN Red List re-assessment for the sub-species. The assessment was prepared by a working group comprising Kenneth /Uiseb, Holger Kolberg, Jeff Muntifering, Sarah King and myself and will be published during 2018 (Gosling, L.M., Muntifering, J., Kolberg, H., Uiseb, K. & King, S.R.B. 2018. Equus zebra ssp. hartmannae. The IUCN Red List of Threatened Species 2018. In press.). The Mountain Zebra Project aims to help support the conservation of the sub-species in Namibia and to study population processes using an individual-based approach. Hartmann’s mountain zebra is a protected species in Namibia and of global conservation importance (Novellie et al, 2002 & 2008; Gosling et al, 2018; IUCN Red List Category: Vulnerable, A3bcd (IUCN version 3.1) and, while locally numerous, they are vulnerable under drought conditions, particularly where seasonal movement is restricted by fences and where they share their range with livestock. -

Equus Zebra Hartmannae – Hartmann’S Mountain Zebra
Equus zebra hartmannae – Hartmann’s Mountain Zebra found no evidence to regard the two taxa as anything more than different populations of a single species. They concluded that the Cape Mountain Zebra and Hartmann’s Mountain Zebra should remain subspecies. Therefore, no taxonomic changes since 2004 have been made. Assessment Rationale Both the South African and Namibian Hartmann’s Mountain Zebra populations have been increasing. For example, over three generations (1980–2015), the subpopulation at Goegap Nature Reserve has increased by 6.2% / year (from 6 to 69 individuals). Similarly, the Cliff and Suretha Dorse subpopulation on Tswalu Kalahari Private Game Reserve has increased from 65 in 2005 to 203 in 2014; and that on Augrabies Falls National Park has increased from 8 in Regional Red List status (2016) Vulnerable D1† 1996 to 208 in 2016. However, subpopulation sizes remain National Red List status (2004) Endangered D small and rely on management to remain viable. In Gondwana Canyon Park, Namibia (adjacent to Ai-Ais/ Reasons for change Genuine change: Richtersveld Transfrontier Park) the Hartmann’s Mountain Population increase Zebra subpopulation has grown at a mean rate of 22% / Global Red List status (2008) Vulnerable C1 year (2005–2012), indicating a healthy source for dispersal and/or translocation into the South African Richtersveld TOPS listing (NEMBA) Vulnerable National Park. From the data available, the observed and CITES listing Appendix II estimated current mature population size within the natural distribution range of South Africa, based on both formally Endemic No and privately protected subpopulations, is at least 592– †Conservation Dependent 724 mature individuals (an underestimate given the lack of comprehensive data on the private sector). -

Zebra: More Than Just Stripes
Zebra: more than just stripes Louw Hoffman Department of Animal Sciences, Stellenbosch University, South Africa SARChI Chair: Meat Science th 9 International Wildlife1 ranching Symposium 12-16 September 2016 Outline • Subspecies • Typical uses • Consumer expectations • Carcass composition & yields • Meat quality • Value addition • Conclusions • Recommendations 2 Introduction • Zebra: 4 sub-species – Equus grevyi (Grevy's zebra) – Equus hartmannae (Hartmann's zebra, Hartmann's mountain zebra) – Equus zebra (Cape mountain zebra, Mountain zebra) – Equus quagga (Plains zebra) 3 Equus grevyi (Grevy's zebra) Source: www.nature.ca4 Equus grevyi (Grevy's zebra) 5 Equus hartmannae (Hartmann's mountain zebra) Source: www.flickr.com6 Equus hartmannae (Hartmann's mountain zebra) 7 Equus zebra (Cape mountain zebra) 8 Equus zebra (Cape mountain zebra) 9 Equus quagga (Plains zebra) Source: commons.Wikimedia.org10 Equus quagga (Plains zebra) 11 12 Typical uses of zebra • Historical & modern 13 Uses Skin 33-37% of the total value 9% of live weight 14 Uses 15 Potential as meat • Biology – Breed well – Stallion & harem of mares – Stallion may kill foals • Harvest – Plains game – easy – Found in large units, economically viable • Diseases – Does not get Foot & Mouth Disease 16 Consumer expectations • Meat should be: – Healthy – Wholesome – Produced ethically – Contain no additives – Produced sustainably – Processable/value addition 17 Somes it up! 18 The Zebra • Harvested in field – Boma – From helicopter – From specialised motor vehicle • Due to value of -

Cape Mountain Zebra (Equus Zebra Zebra) Habitat Use and Diet in the Bontebok National Park, South Africa
Cape mountain zebra (Equus zebra zebra) habitat use and diet in the Bontebok National Park, South Africa by Taniia Strauss Submitted in fulfilment of the requirements for the degree of Masters of Technology to be awarded at the Nelson Mandela Metropolitan University January 2015 Supervisor: Prof. L.H. Watson DECLARATION I, Taniia Strauss (student number 20106975), hereby declare that the thesis for Degree of Masters of Technology to be awarded is my own work, and has not previously been submitted for assessment or completion of any postgraduate qualification to another University or for another qualification. _________________ Taniia Strauss Official use: In accordance with Rule G4.6.3, 4.6.3 A treatise/dissertation/thesis must be accompanied by a written declaration on the part of the candidate to the effect that it is his/her own work and that it has not previously been submitted for assessment to another University or for another qualification. However, material from publications by the candidate may be embodied in a treatise/dissertation/thesis. I ABSTRACT Cape mountain zebra habitat utilization and diet in the dystrophic fynbos habitat of the Bontebok National Park was found to be highly selective. Mountain zebra concentrated in specific sectors and habitat types in the park on a seasonal basis, preferring recently burnt habitat with a veld age younger than one year in all seasons, except during the warm, dry summer. Proteoid Fynbos with a veld age between one and five years was preferred in the cool winter, while Drainage Lines and the fringes of Inland Pans were preferred during summer. -

Preventing Hybridization of Cape Mountain Zebra
PREVENTING HYBRIDIZATION OF CAPE MOUNTAIN ZEBRA There are three types of zebra in the Western Cape but only one, the Cape mountain zebra (CMZ, PHOTO: Martin Harvey PHOTO: Equus zebra zebra), occurs here naturally. Cape mountain zebra Hartmann’s zebra (Equus zebra hartmannae) and CAN YOU SPOT plains zebra (Equus quagga burchelli) were introduced THE DIFFERENCE? into the province. Some of these animals escaped from their points of introduction and now roam freely across the landscape. Both plains and Hartmann’s zebra can mate with CMZ. Foals resulting from a CMZ-Hartmann’s cross will The quagga, Equus quagga probably be fertile, resulting in contamination of both quagga, a subspecies of the the CMZ and Hartmann’s gene pools. Although CMZ- plains zebra, used to occur plains foals will likely be infertile, such crosses are wasted Cape mountain zebra in the Western Cape but reproductive opportunities for both species. Zebra can is now extinct. also interbreed with donkeys and horses. Hartmann’s zebra PHOTO: Kevin Shaw Kevin PHOTO: Trail camera image of a cross between a wild Horse-CMZ hybrid, Bredasdorp area horse and a Hartmann’s zebra (Namibia) Plains zebra Familiarize yourself with the WHAT YOU CAN DO TO HELP differences between the different zebra types, using this pamphlet as a If you keep zebra on your property or want guide (see overleaf). But note that although to introduce zebra, please make sure that some hybrids look obviously different from CMZ, not all you have a permit from CapeNature to do hybrids can be distinguished on the basis of physical features so. -

Cape Mountain Zebra Was Almost Extinct As a Result of Hunting and Competition for Grazing with Farm Stock
CapeCape MountainMountain ZebraZebra Equus zebra zebra Conservation needs In the 1930s the Cape mountain zebra was almost extinct as a result of hunting and competition for grazing with farm stock. Conservation initiatives such as the formation of Mountain Zebra National Park in 1937 led to a steady increase in numbers. Cape mountain zebras are still rare and classed as endangered (IUCN Endangered; CITES Appendix I). The conservation priority is to build up numbers as quickly as possible to ensure long-term survival.. -
MZNP Mammal List 2013
Mammals checklist of Mountain Zebra National Park Order Insectivora Order Family Erinaceidae Family South African Suid-Afrikaanse Erinaceus frontalis Porcupine hedgehog krimpvarkie Family Order Primates Ground squirrel Vervet monkey Blouaap Cercopithecus pygerythrus Chacma baboon Kaapse bobbejaan Papio ursinus Family Order Carnivora Springhare Family Protelidae Aardwolf Aardwolf Proteles cristatus Family African pygmy mouse Family Hyaenidae Brown hyena Strandwolf Parahyaena brunnea Order Family Family Felidae Cape mountain zebra Cape wild cat Vaalboskat Felis lybica Burchell's zebra Black-footed cat Kleingekoldekat Felis nigripes Caracal Rooikat Caracal caracal Family Cheetah Jagluiperd Acinonyx jubatus Black rhinoceros Lion Leeu Panthera leo Order Family Canidae Family Bat-eared fox Bakoorjakkals Otocyon megalotis Bushpig Cape fox Silwervos Vulpes chama Blackbacked jackal Rooi jakkals Canis mesomelas Family Eland Family Mustelidae Kudu Cape clawless otter Groototter Aonyx capensis Mountain reedbuck Striped polecat Stinkmuishond Ictonyx striatus Red hartebeest African striped weasel Slangmuishond Poecilogale albinucha Black wildebeest Honey badger Ratel Mellivora capensis Cape buffalo Bushbuck Family Viverridae Blesbuck Small-spotted genet Kleinkulmuskoljaatkat Genetta genetta Springbuck Cape grey mongoose Kleingrysmuishond Galerella pulverulenta Grey duiker Water mongoose Kommetjiegatmuishond Atilax paludinosus Gemsbuck Yellow mongoose Geelmuishond Cynctis penicillata Steenbuck Suricate / Meerkat Stokstertmeerkat Suricata suricatta Klipspringer -
Camdeboo-Mountain Zebra National Park Corridor: Opportunities for Conservation and Socio-Economic Development
Camdeboo-Mountain Zebra National Park Corridor: Opportunities for Conservation and Socio-Economic Development Matthew Norval Abstract—The Wilderness Foundation, in partnership with South The Mountain Zebra National Park Complex is an identi- African National Parks has initiated a two year project in the Karoo; fied Critical Ecosystem Partnership Fund (CEPF) priority The Mountain Zebra-Camdeboo Corridor Project. Through either with the prioritization based on the presence of a range voluntary Contractual National Park or Protected Environment of Red Listed Species (especially the larger charismatic agreements, the project aims to work with, rather than displace, threatened mammals), the potential importance as a climate current conservation-compatible land-use practices such as eco- change adaptation corridor, and to a lesser extent for the tourism, livestock grazing and other sustainable resource use. Less delivery of ecosystem goods and services. The area forms a formal conservation options, such as biodiversity agreements, will large part of a corridor known as the Amathole-Sneeuberg also be offered to landowners. These agreements provide benefits Montane Belt, and now also contains the recently identified to landowners in terms of protecting the environment as well as Sneeuberg Centre of Endemism. contributing towards the conservation of threatened wildlife spe- The project aims to capitalise on the opportunities created cies. Protection contributes to safeguarding the recently-identified through the emergence of a significant private conservation Sneeuberg Centre of Endemism, part of the Amathole-Sneeuberg land holding in the area with land use compatible for formal Montane Belt. The project will stimulate conservation-friendly inclusion as contractual national parks. If this opportunity economic development in the region while protecting it from inap- is not pursued immediately there is a danger that the con- propriate development. -
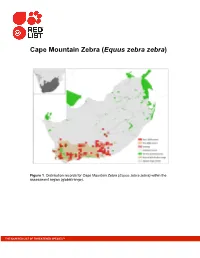
Cape Mountain Zebra (Equus Zebra Zebra)
Cape Mountain Zebra (Equus zebra zebra) Figure 1. Distribution records for Cape Mountain Zebra (Equus zebra zebra) within the assessment region (global range). THE IUCN RED LIST OF THREATENED SPECIES™ Table 1. Countries of occurrence within southern Africa Country Presence Origin Botswana Absent — Lesotho Absent — Mozambique Absent — Namibia Absent — South Africa: Eastern Cape Extant Native South Africa: Western Cape Extant Native South Africa: Free State Extant Introduced South Africa: Northern Cape Extant Native & Introduced Swaziland Absent — Zimbabwe Absent — Figure 2. Historical distribution of Cape Mountain Zebra (Equus zebra zebra) within the assessment region: background colours represent biomes (source: Boshoff et al. 2015). Table 2. Subpopulation numbers of Cape Mountain Zebra (Equus zebra zebra) aggregated by province, 2009 to 2014/15. Inside No. of Sub- Sub- Mature Mature natural reserves/ population population 55% 75% Province Type distribution properties total total range (2009, 2014) (2009) (2013-2015) Eastern Cape Formally protected Yes 6 1,022 1,903 1,047 11,427 Private Yes 10, 26 319 868 477 651 Northern Cape Formally protected Yes 2 34 35 19 26 Western Cape Formally protected Yes 10 821 1,179 648 884 Private Yes 23, 27 473 628 345 471 Free State Formally protected No 1 93 112 62 84 Private No 1 Unknown 8 4 6 Northern Cape Private No Unknown Unknown Unknown Total formally protected (eligible) 18 3,117 1,714 2,338 Total privately protected (eligible) 53 1,496 823 1,122 Extrapolated total eligible (81%) 53 1,212 666 909 Grand total 55 4,733 2,603 3,550 Total eligible adjusted 53 4,329 2,381 3,247 Table 3.