Seed Mucilage Evolution
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Cruciferae) – Mustard Family
BRASSICACEAE (CRUCIFERAE) – MUSTARD FAMILY Plant: herbs mostly, annual to perennial, sometimes shrubs; sap sometimes peppery Stem: Root: Leaves: mostly simple but sometimes pinnately divided; alternate, rarely opposite or whorled; no stipules Flowers: mostly perfect, mostly regular (actinomorphic); 4 sepals, 4 petals often forming a cross; 6 stamens with usually 2 outer ones shorter than the inner 4; ovary superior, mostly 2 fused carpels, 1 to many ovules, 1 pistil Fruit: seed pods, often used in classification, many are slender and long (Silique), some broad (Silicle) – see morphology slide Other: a large family, many garden plants such as turnip, radish, and cabbage, also some spices; often termed the Cruciferae family; Dicotyledons Group Genera: 350+ genera; 40+ locally WARNING – family descriptions are only a layman’s guide and should not be used as definitive Flower Morphology in the Brassicaceae (Mustard Family) - flower with 4 sepals, 4 petals (often like a cross, sometimes split or lobed), commonly small, often white or yellow, distinctive fruiting structures often important for ID 2 types of fruiting pods: in addition, fruits may be circular, flattened or angled in cross-section Silicle - (usually <2.5x long as wide), 2-valved with septum (replum) Silique - (usually >2.5x long as wide), 2- valved with septum (replum) Flowers, Many Genera BRASSICACEAE (CRUCIFERAE) – MUSTARD FAMILY Sanddune [Western] Wallflower; Erysimum capitatum (Douglas ex Hook.) Greene var. capitatum Wormseed Wallflower [Mustard]; Erysimum cheiranthoides L. (Introduced) Spreading Wallflower [Treacle Mustard]; Erysimum repandum L. (Introduced) Dame’s Rocket [Dame’s Violet]; Hesperis matronalis L. (Introduced) Purple [Violet] Rocket; Iodanthus pinnatifidus (Michx.) Steud. Michaux's Gladecress; Leavenworthia uniflora (Michx.) Britton [Cow; Field] Cress [Peppergrass]; Lepidium campestre L.) Ait. -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Planting Planner
what seeds to sow BRIZA MEDIA Planting ‘GOLDEN BEE’ I love quaking grasses and this selection has a golden tint to the planner seedheads as they age. SLP†. Height (H) 60cm. Sow (S) Best The best seeds to sown in situ in spring. Period of interest (P) sow for 2015 May – September. Winter is the perfect time for studying seed catalogues and planning what to grow. Leading designer Sarah Price gives us a sneak peek of her list WORDS SARAH PRICE PERSICARIA ORIENTALIS Definitely a plant for the back of the border, this tall, multi-branched annual has dramatic, long, deep pink, drooping plumes. Popular in the Victorian era. HA. H 1.8m. S Best sown fresh in autumn. CLOCKWISE FROM TOP: JASON INGRAM X2; STEFFEN HAUSER / GARDEN WORLD IMAGES; NICHOLAS APPLEBY / GARDEN WORLD IMAGES / GARDEN WORLD APPLEBY NICHOLAS IMAGES; / GARDEN WORLD HAUSER X2; STEFFEN INGRAM JASON TOP: FROM CLOCKWISE Can also be sown in early spring. his year my cash-strapped local authority P Late summer to autumn. Twas flooded with unlikely letters of love and appreciation after it set verges and roundabouts alive with a blaze of LATHYRUS annual colour instead of the usual regimented bedding. VERNUS ‘FILIGREE’ It’s proof that broadcasting a packet, or in this case Low-growing, bucketfuls, of mixed seed can have mood-enhancing bushy, hardy effects. It’s not all about live-fast-die-young annuals. perennial sweet pea, Many of my favourite plants are hardy biennials, which invaluable for its take two years to flower, set seed and die, while others are early, purple-blue hardy herbaceous perennials that will live for years. -

State of New York City's Plants 2018
STATE OF NEW YORK CITY’S PLANTS 2018 Daniel Atha & Brian Boom © 2018 The New York Botanical Garden All rights reserved ISBN 978-0-89327-955-4 Center for Conservation Strategy The New York Botanical Garden 2900 Southern Boulevard Bronx, NY 10458 All photos NYBG staff Citation: Atha, D. and B. Boom. 2018. State of New York City’s Plants 2018. Center for Conservation Strategy. The New York Botanical Garden, Bronx, NY. 132 pp. STATE OF NEW YORK CITY’S PLANTS 2018 4 EXECUTIVE SUMMARY 6 INTRODUCTION 10 DOCUMENTING THE CITY’S PLANTS 10 The Flora of New York City 11 Rare Species 14 Focus on Specific Area 16 Botanical Spectacle: Summer Snow 18 CITIZEN SCIENCE 20 THREATS TO THE CITY’S PLANTS 24 NEW YORK STATE PROHIBITED AND REGULATED INVASIVE SPECIES FOUND IN NEW YORK CITY 26 LOOKING AHEAD 27 CONTRIBUTORS AND ACKNOWLEGMENTS 30 LITERATURE CITED 31 APPENDIX Checklist of the Spontaneous Vascular Plants of New York City 32 Ferns and Fern Allies 35 Gymnosperms 36 Nymphaeales and Magnoliids 37 Monocots 67 Dicots 3 EXECUTIVE SUMMARY This report, State of New York City’s Plants 2018, is the first rankings of rare, threatened, endangered, and extinct species of what is envisioned by the Center for Conservation Strategy known from New York City, and based on this compilation of The New York Botanical Garden as annual updates thirteen percent of the City’s flora is imperiled or extinct in New summarizing the status of the spontaneous plant species of the York City. five boroughs of New York City. This year’s report deals with the City’s vascular plants (ferns and fern allies, gymnosperms, We have begun the process of assessing conservation status and flowering plants), but in the future it is planned to phase in at the local level for all species. -

Phytochemical Profile and Antioxidant Activity of the Aerial Part Extracts from Matthiola Incana Subsp
DOI: 10.1002/cbdv.202100167 FULL PAPER Phytochemical Profile and Antioxidant Activity of the Aerial Part Extracts from Matthiola incana subsp. rupestris and subsp. pulchella (Brassicaceae) Endemic to Sicily Natalizia Miceli,*a Emilia Cavò,a, b Vivienne Spadaro,c Francesco Maria Raimondo,d Salvatore Ragusa,e Francesco Cacciola,f Yassine Oulad El Majdoub,a Katia Arena,a Luigi Mondello,a, g, h, i Concetta Condurso,j Fabrizio Cincotta,j and Maria Fernanda Tavianoa a Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Viale Palatucci, 98168 Messina, Italy, e-mail: [email protected] b Foundation ‘Prof. Antonio Imbesi’, University of Messina, Piazza Pugliatti 1, 98122 Messina, Italy, c Department STEBICEF/Section of Botany, Anthropology and Zoology, University of Palermo, Via Archirafi 38, 90123 Palermo, Italy, d PLANTA/Research, Documentation and Training Center, Via Serraglio Vecchio 28, 90123 Palermo, Italy, e Department of Health Sciences, University ‘Magna Graecia’ of Catanzaro, Viale Europa, Località Germaneto, 88100 Catanzaro, Italy, f Department of Biomedical, Dental, Morphological and Functional Imaging Sciences, University of Messina, Via Consolare Valeria, 98125 Messina, Italy, g Chromaleont s.r.l., c/o Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Viale Palatucci, 98168 Messina, Italy, h BeSep s.r.l., c/o Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Viale Palatucci, 98168 Messina, Italy, i Department of Sciences and Technologies for Human and Environment, University Campus Bio-Medico of Rome, via Àlvaro del Portillo 21, 00128 Rome, Italy, j Department of Veterinary Sciences, University of Messina, Viale Palatucci, 98168 Messina, Italy. -

Conserving Europe's Threatened Plants
Conserving Europe’s threatened plants Progress towards Target 8 of the Global Strategy for Plant Conservation Conserving Europe’s threatened plants Progress towards Target 8 of the Global Strategy for Plant Conservation By Suzanne Sharrock and Meirion Jones May 2009 Recommended citation: Sharrock, S. and Jones, M., 2009. Conserving Europe’s threatened plants: Progress towards Target 8 of the Global Strategy for Plant Conservation Botanic Gardens Conservation International, Richmond, UK ISBN 978-1-905164-30-1 Published by Botanic Gardens Conservation International Descanso House, 199 Kew Road, Richmond, Surrey, TW9 3BW, UK Design: John Morgan, [email protected] Acknowledgements The work of establishing a consolidated list of threatened Photo credits European plants was first initiated by Hugh Synge who developed the original database on which this report is based. All images are credited to BGCI with the exceptions of: We are most grateful to Hugh for providing this database to page 5, Nikos Krigas; page 8. Christophe Libert; page 10, BGCI and advising on further development of the list. The Pawel Kos; page 12 (upper), Nikos Krigas; page 14: James exacting task of inputting data from national Red Lists was Hitchmough; page 16 (lower), Jože Bavcon; page 17 (upper), carried out by Chris Cockel and without his dedicated work, the Nkos Krigas; page 20 (upper), Anca Sarbu; page 21, Nikos list would not have been completed. Thank you for your efforts Krigas; page 22 (upper) Simon Williams; page 22 (lower), RBG Chris. We are grateful to all the members of the European Kew; page 23 (upper), Jo Packet; page 23 (lower), Sandrine Botanic Gardens Consortium and other colleagues from Europe Godefroid; page 24 (upper) Jože Bavcon; page 24 (lower), Frank who provided essential advice, guidance and supplementary Scumacher; page 25 (upper) Michael Burkart; page 25, (lower) information on the species included in the database. -

Plant Life MagillS Encyclopedia of Science
MAGILLS ENCYCLOPEDIA OF SCIENCE PLANT LIFE MAGILLS ENCYCLOPEDIA OF SCIENCE PLANT LIFE Volume 4 Sustainable Forestry–Zygomycetes Indexes Editor Bryan D. Ness, Ph.D. Pacific Union College, Department of Biology Project Editor Christina J. Moose Salem Press, Inc. Pasadena, California Hackensack, New Jersey Editor in Chief: Dawn P. Dawson Managing Editor: Christina J. Moose Photograph Editor: Philip Bader Manuscript Editor: Elizabeth Ferry Slocum Production Editor: Joyce I. Buchea Assistant Editor: Andrea E. Miller Page Design and Graphics: James Hutson Research Supervisor: Jeffry Jensen Layout: William Zimmerman Acquisitions Editor: Mark Rehn Illustrator: Kimberly L. Dawson Kurnizki Copyright © 2003, by Salem Press, Inc. All rights in this book are reserved. No part of this work may be used or reproduced in any manner what- soever or transmitted in any form or by any means, electronic or mechanical, including photocopy,recording, or any information storage and retrieval system, without written permission from the copyright owner except in the case of brief quotations embodied in critical articles and reviews. For information address the publisher, Salem Press, Inc., P.O. Box 50062, Pasadena, California 91115. Some of the updated and revised essays in this work originally appeared in Magill’s Survey of Science: Life Science (1991), Magill’s Survey of Science: Life Science, Supplement (1998), Natural Resources (1998), Encyclopedia of Genetics (1999), Encyclopedia of Environmental Issues (2000), World Geography (2001), and Earth Science (2001). ∞ The paper used in these volumes conforms to the American National Standard for Permanence of Paper for Printed Library Materials, Z39.48-1992 (R1997). Library of Congress Cataloging-in-Publication Data Magill’s encyclopedia of science : plant life / edited by Bryan D. -

Seed Germination and Genetic Structure of Two Salvia Species In
Seed germination and genetic structure of two Salvia species in response to environmental variables among phytogeographic regions in Jordan (Part I) and Phylogeny of the pan-tropical family Marantaceae (Part II). Dissertation Zur Erlangung des akademischen Grades Doctor rerum naturalium (Dr. rer. nat) Vorgelegt der Naturwissenschaftlichen Fakultät I Biowissenschaften der Martin-Luther-Universität Halle-Wittenberg Von Herrn Mohammad Mufleh Al-Gharaibeh Geb. am: 18.08.1979 in: Irbid-Jordan Gutachter/in 1. Prof. Dr. Isabell Hensen 2. Prof. Dr. Martin Roeser 3. Prof. Dr. Regina Classen-Bockhof Halle (Saale), den 10.01.2017 Copyright notice Chapters 2 to 4 have been either published in or submitted to international journals or are in preparation for publication. Copyrights are with the authors. Just the publishers and authors have the right for publishing and using the presented material. Therefore, reprint of the presented material requires the publishers’ and authors’ permissions. “Four years ago I started this project as a PhD project, but it turned out to be a long battle to achieve victory and dreams. This dissertation is the culmination of this long process, where the definition of “Weekend” has been deleted from my dictionary. It cannot express the long days spent in analyzing sequences and data, battling shoulder to shoulder with my ex- computer (RIP), R-studio, BioEdite and Microsoft Words, the joy for the synthesis, the hope for good results and the sadness and tiredness with each attempt to add more taxa and analyses.” “At the end, no phrase can describe my happiness when I saw the whole dissertation is printed out.” CONTENTS | 4 Table of Contents Summary .......................................................................................................................................... -

I Pathotypes of Plasmodiophora Brassicae from Clubroot Resistant
Pathotypes of Plasmodiophora brassicae from clubroot resistant canola and assessment of amisulbrom for clubroot control By Keisha Hollman A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science In Plant Science Department of Agricultural, Food and Nutritional Science University of Alberta ©Keisha Hollman, 2021 i Abstract Clubroot, caused by Plasmodiophora brassicae Woronin, is an important soilborne disease of canola (oilseed rape; Brassica napus L.). In Canada, clubroot management relies heavily on the planting of resistant cultivars, but since 2013, resistance has been broken in an increasing number of fields. Prior to the introduction of resistance, P. brassicae pathotype 3H, as defined on the Canadian Clubroot Differential (CCD) set, was predominant in Alberta. In testing of pathogen collections from 2014-2016, however, pathotype 3A was most common, indicating rapid shifts in the pathogen population. Up-to-date knowledge of pathotype composition is important for effective resistance breeding and stewardship. Furthermore, strategies to supplement resistance, such as the application of fungicides, may also contribute to sustainable clubroot management. In this thesis, isolates of P. brassicae were obtained from 166 canola crops in Alberta, Saskatchewan and Manitoba, and evaluated for pathotype designation on the CCD set and the differentials of Somé et al. Seventeen pathotypes were detected on the CCD set, including the previously reported pathotypes 3A, 3D, 3H, 5L, 5X, 8E, 8N and 8P, plus the novel pathotypes 2C, 6D, 8D, 9A, 9B, 9C, 11A, 13A and 13B. Five pathotypes were identified on the hosts of Somé et al. including P1, P2, P3, P4 and P5, with P4 and P5 reported here from Canada for the first time. -

Albugo S.Str. (Albuginales; Oomycota) Is Not Restricted to Brassicales but Also Present on Fabales
Org Divers Evol (2011) 11:193–199 DOI 10.1007/s13127-011-0043-5 ORIGINAL ARTICLE A new perspective on the evolution of white blister rusts: Albugo s.str. (Albuginales; Oomycota) is not restricted to Brassicales but also present on Fabales Young-Joon Choi & Marco Thines & Hyeon-Dong Shin Received: 21 July 2010 /Accepted: 28 February 2011 /Published online: 13 March 2011 # Gesellschaft für Biologische Systematik 2011 Abstract For almost all groups of pathogens, unusual and Resedaceae in the order Brassicales. In the present study, rare host species have been reported. Often, such associa- molecular phylogenetic analysis of cox2 mtDNA sequences tions are based on single or few collections only, which are and morphological investigations on an original specimen frequently hard to access. Many of them later prove to be confirmed the occurrence of a member of Albugo on due to misidentification of the host, the pathogen, or both. Fabaceae hosts, with the characteristic thin wall of the Therefore, such reports are often disregarded, or treated secondary sporangia, which is almost uniform in thickness. anecdotally in taxonomic and phylogenetic studies, regard- In phylogenetic analyses the species results as embedded less of their potential importance to unravelling the within Albugo s.str. Therefore, it is concluded that the evolution of the entire group. Concerning oomycete natural host range of Albugo s.str. extends from Brassicales biotrophs there are several reports of unusual and rare to Fabales via host jumping. Our results underscore that hosts for hardly known pathogens. In the order Fabales, for unrevised reports of pathogens from unusual hosts should example, a single species of Albugo, A. -

Flora Mediterranea 26
FLORA MEDITERRANEA 26 Published under the auspices of OPTIMA by the Herbarium Mediterraneum Panormitanum Palermo – 2016 FLORA MEDITERRANEA Edited on behalf of the International Foundation pro Herbario Mediterraneo by Francesco M. Raimondo, Werner Greuter & Gianniantonio Domina Editorial board G. Domina (Palermo), F. Garbari (Pisa), W. Greuter (Berlin), S. L. Jury (Reading), G. Kamari (Patras), P. Mazzola (Palermo), S. Pignatti (Roma), F. M. Raimondo (Palermo), C. Salmeri (Palermo), B. Valdés (Sevilla), G. Venturella (Palermo). Advisory Committee P. V. Arrigoni (Firenze) P. Küpfer (Neuchatel) H. M. Burdet (Genève) J. Mathez (Montpellier) A. Carapezza (Palermo) G. Moggi (Firenze) C. D. K. Cook (Zurich) E. Nardi (Firenze) R. Courtecuisse (Lille) P. L. Nimis (Trieste) V. Demoulin (Liège) D. Phitos (Patras) F. Ehrendorfer (Wien) L. Poldini (Trieste) M. Erben (Munchen) R. M. Ros Espín (Murcia) G. Giaccone (Catania) A. Strid (Copenhagen) V. H. Heywood (Reading) B. Zimmer (Berlin) Editorial Office Editorial assistance: A. M. Mannino Editorial secretariat: V. Spadaro & P. Campisi Layout & Tecnical editing: E. Di Gristina & F. La Sorte Design: V. Magro & L. C. Raimondo Redazione di "Flora Mediterranea" Herbarium Mediterraneum Panormitanum, Università di Palermo Via Lincoln, 2 I-90133 Palermo, Italy [email protected] Printed by Luxograph s.r.l., Piazza Bartolomeo da Messina, 2/E - Palermo Registration at Tribunale di Palermo, no. 27 of 12 July 1991 ISSN: 1120-4052 printed, 2240-4538 online DOI: 10.7320/FlMedit26.001 Copyright © by International Foundation pro Herbario Mediterraneo, Palermo Contents V. Hugonnot & L. Chavoutier: A modern record of one of the rarest European mosses, Ptychomitrium incurvum (Ptychomitriaceae), in Eastern Pyrenees, France . 5 P. Chène, M. -
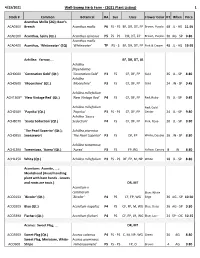
WSHF Catalog
4/23/2021 Well-Sweep Herb Farm - (2021 Plant Listing) 1 Stock # Common Botanical HA Sun Uses Flower ColorHT When Price Acanthus Mollis (2Q); Bear's ACA030X Breech Acanthus mollis P6 FS - PS BF, DR, DT, FP Brown, Purple- 48 JL - AG 11.95 ACA010X Acanthus, Spiny (Qt.) Acanthus spinosus P5 FS - PS DR, DT, FP Brown, Purple- 30 AG - SP 9.80 Acanthus mollis ACA040X Acanthus, `Whitewater' (2Q) `Whitewater' TP PS - S BF, DR, DT, FP Pink & Cream 48 JL - AG 19.95 Achillea: Yarrow, ... BF, DR, DT, LB Achillea filipendulina ACH000X `Coronation Gold' (Qt.) `Coronation Gold' P3 FS CF, DF, FP Gold 36 JL - SP 8.80 Achillea ACH050X `Moonshine' (Qt.) `Moonshine' P3 FS CF, DF, FP Gold 24 JN - SP 9.45 Achillea millefolium ACH130X* `New Vintage Red' (Qt.) `New Vintage Red' P4 FS CF, DF, FP Red, Ruby- 15 JL - SP 9.45 Achillea millefolium Red; Gold ACH250X `Paprika' (Qt.) `Paprika' P3 FS - PS CF, DF, FP Center 24 JL - SP 9.80 Achillea `Saucy ACH807X `Saucy Seduction' (Qt.) Seduction' P4 FS CF, DF, FP Pink, Rose- 20 JL - SP 9.80 `The Pearl Superior' (Qt.); Achillea ptarmica ACH095X Sneezewort `The Pearl Superior' P3 FS DF, FP White; Double 16 JN - SP 8.80 Achillea tomentosa ACH120X Tomentosa, `Aurea' (Qt.) `Aurea' P3 FS FP, RG Yellow, Canary- 8 JN 8.80 ACH125X White (Qt.) Achillea millefolium P3 FS - PS DF, FP, M, NP White 18 JL - SP 8.80 Aconitum: Aconite, ... ; Monkshood (Avoid handling plant with bare hands - Leaves and roots are toxic.) DR, MT Aconitum x cammarum Blue; White ACO015X `Bicolor' (Qt.) `Bicolor' P4 PS CF, FP, WG Edge 36 AG - SP 10.50 ACO020X Blue (Qt.) Aconitum napellus P4 PS CF, FP, M, WG Blue, Deep- 36 AG - SP 9.80 ACO339X Fischeri (Qt.) Aconitum fischeri P4 PS CF, FP, LB, WG Blue, Lav.- 24 SP - OC 10.15 Acorus: Sweet Flag, ..