ENHANCING SCIENCE EDUCATION THROUGH TEACHING ORIGINS in SCIENCE CLASSES by David A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Early Eocene Primates from Gujarat, India
ARTICLE IN PRESS Journal of Human Evolution xxx (2009) 1–39 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Early Eocene Primates from Gujarat, India Kenneth D. Rose a,*, Rajendra S. Rana b, Ashok Sahni c, Kishor Kumar d, Pieter Missiaen e, Lachham Singh b, Thierry Smith f a Johns Hopkins University School of Medicine, Baltimore, Maryland 21205, USA b H.N.B. Garhwal University, Srinagar 246175, Uttarakhand, India c Panjab University, Chandigarh 160014, India d Wadia Institute of Himalayan Geology, Dehradun 248001, Uttarakhand, India e University of Ghent, B-9000 Ghent, Belgium f Royal Belgian Institute of Natural Sciences, B-1000 Brussels, Belgium article info abstract Article history: The oldest euprimates known from India come from the Early Eocene Cambay Formation at Vastan Mine Received 24 June 2008 in Gujarat. An Ypresian (early Cuisian) age of w53 Ma (based on foraminifera) indicates that these Accepted 8 January 2009 primates were roughly contemporary with, or perhaps predated, the India-Asia collision. Here we present new euprimate fossils from Vastan Mine, including teeth, jaws, and referred postcrania of the Keywords: adapoids Marcgodinotius indicus and Asiadapis cambayensis. They are placed in the new subfamily Eocene Asiadapinae (family Notharctidae), which is most similar to primitive European Cercamoniinae such as India Donrussellia and Protoadapis. Asiadapines were small primates in the size range of extant smaller Notharctidae Adapoidea bushbabies. Despite their generally very plesiomorphic morphology, asiadapines also share a few derived Omomyidae dental traits with sivaladapids, suggesting a possible relationship to these endemic Asian adapoids. In Eosimiidae addition to the adapoids, a new species of the omomyid Vastanomys is described. -

Anthroquest the Newsletter of the Leakey Foundation
no. 35 Spring/Summer 2017 AnthroQuest The Newsletter of The Leakey Foundation Leakey Grantee Finds Oldest Primates of India THIERRY SMITH, LEAKEY FOUNDATION GRANTEE, ROYAL BELGIAN INSTITUTE OF NATURAL SCIENCES n 2004, Professor Kenneth D. Rose from John Hopkins I University invited me to lead a small team of paleontologists planning to explore lignite (brown coal) mines in India. The goal was to prospect for Paleocene-Eocene deposits (66-34 million years ago), which could yield ancient terrestrial mammals. Ken thought the screen-washing technique we used to find small fossil mammals in Wyoming would work well in India. I went to Mumbai with a student, Pieter Missiaen, and we travelled about 250 mile north to the Vastan mine in Thierry Smith screenwashing sediments for small remains of earliest Indian primates Gujarat to meet our Indian colleagues Photo: Annelise Folie for the first time. I remember well that on the second day in the mine, it was a mammal incisor the size of a human million years old). We published this hot, and I was nervous because we were incisor. It was the beginning of our jaw with four teeth under the new name six persons sharing only two hammers Indian adventure! Asiadapis cambayensis, and today this to look for fossils in a sticky clay layer. specimen is the reference of the family This deposit was so rich that we Asiadapidae. I suggested to my colleague, Professor collected all the matrix and screen- Rajendra Singh Rana, that he ask a washed it completely. Each sieve The Vastan mine was a paradise for miner to remove the lignite layer above contained small mammal teeth or paleontologists. -

Effects of Additional Postcranial Data on the Topologies of Early Eocene Primate
Effects of Additional Postcranial Data on the Topologies of Early Eocene Primate Phylogenies Julia Stone ANT 679HB Special Honors in the Department of Anthropology The University of Texas at Austin May 2021 Liza J. Shapiro Department of Anthropology Supervising Professor E. Christopher Kirk Department of Anthropology Second Reader Acknowledgements This project would not have been completed without the help of my thesis supervisor Dr. Liza Shapiro. I was inspired to start the project after taking her courses in Primate Evolution and Primate Anatomy. She also met with me many times throughout this year to discuss the trajectory of the project as well as discuss papers that became the foundation of the thesis. She assisted me through technology issues, unresolved trees, and many drafts, and she provided invaluable guidance throughout this process. I also want to thank Dr. Christopher Kirk for being my second reader and providing extremely helpful insight into how to write about phylogenetic results. He also helped me to be more specific and accurate in all of the sections of the thesis. I learned a lot about writing theses in general due to his comments on my drafts. I also truly appreciate the help of Ben Rodwell, a graduate student at the University of Texas at Austin. He provided excellent help and support regarding the Mesquite and TNT software used in this project. ii Effects of Additional Postcranial Data on the Topologies of Early Eocene Primate Phylogenies by Julia Stone, BA The University of Texas at Austin SUPERVISOR: Liza J. Shapiro There are multiple hypotheses regarding the locomotor behaviors of the last common ancestor to primates and which fossil primates best represent that ancestor. -
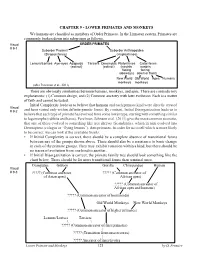
CHAPTER 9 - LOWER PRIMATES and MONKEYS We Humans Are Classified As Members of Order Primates
CHAPTER 9 - LOWER PRIMATES AND MONKEYS We humans are classified as members of Order Primates. In the Linnaean system, Primates are commonly broken down into subgroups as follows. Visual ORDER PRIMATES # 9-1 Suborder Prosimii Suborder Anthropoidea (Strepsirrhines) (Haplorrhines) Lemurs/Lorises Aye-ayes Adapoids Tarsiers Omomyids Platyrrhines Catarrhines (extinct) (extinct) (nostrils nostrils facing facing sideways) down or front) New World Old World Apes Humans monkeys monkeys (after Perelman et al., 2011) There are obviously similarities between humans, monkeys, and apes. There are contradictory explanations: (1) Common design, and (2) Common ancestry with later evolution. Each is a matter of faith and cannot be tested. Initial Complexity leads us to believe that humans and each primate kind were directly created Visual # 9-2 and have varied only within definite genetic limits. By contrast, Initial Disorganization leads us to believe that each type of primate has evolved from some lower type, starting with something similar to lagomorphs (rabbits and hares). Perelman, Johnson et al. (2011) give the most common scenario, that one of these evolved to something like tree shrews (Scandentia), which in turn evolved into Dermoptera (colugos or “flying lemurs”), then primates. In order for us to tell which is more likely to be correct, we can look at the available fossils. • If Initial Complexity is correct, there should be a complete absence of transitional forms between any of the groups shown above. There should also be a resistance to basic change in each of the primate groups. They may exhibit variation within a kind, but there should be no traces of evolution from one kind to another. -

Nous Primats De L'eocè De La Península Ibèrica
Departament de Biologia Animal, de Biologia Vegetal i d’Ecologia Unitat d’Antropologia Biològica Nous primats de l’Eocè de la Península Ibèrica: implicacions filogenètiques i paleobiogeogràfiques Judit Marigó Cortés Tesi Doctoral 2013 Il·lustració de la portada: Óscar Sanisidro Departament de Biologia Animal, de Biologia Vegetal i d’Ecologia Unitat d’Antropologia Biològica Nous primats de l’Eocè de la Península Ibèrica: implicacions filogenètiques i paleobiogeogràfiques Memòria presentada per Judit Marigó Cortés per optar al títol de Doctor en Biologia, programa de doctorat en Biodiversitat del Departament de Biologia Animal, de Biologia Vegetal i d’Ecologia de la Universitat Autònoma de Barcelona, dirigida per: - Dr. Salvador Moyà Solà, ICREA a l’Institut Català de Paleontologia Miquel Crusafont i a la Unitat d’Antropologia Biològica del Departament de Biologia Animal, de Biologia Vegetal i d’Ecologia de la Universitat Autònoma de Barcelona. - Dr. Raef Minwer-Barakat Requena, Institut Català de Paleontologia Miquel Crusafont. Dr. Salvador Moyà Solà Dr. Raef Minwer-Barakat Requena Judit Marigó Cortés TREBALL FINANÇAT PER: - Institut Català de Paleontologia Miquel Crusafont. - Generalitat de Catalunya, Departament d’Innovació, Universitats i Empresa i AGAUR, beca predoctoral 2010FIB176 i beques de mobilitat BE i CTP (2010BE00758, 2011BE100009 i 2011CTP00011), i Grup de Recerca Consolidat "Grup de Paleoprimatologia i Paleontologia Humana PIPH". - Gobierno de España, Ministerio de Ciencia e Innovación y Ministerio de Economía y Competitividad, proyectos "Grandes simios fósiles (Hominoidea) del Mioceno del area Mediterránea: origen, paleobiología y evolución (CGL2008-00325/BTE)” e “Historia Evolutiva de los Primates del Paleógeno y Neógeno de la Península Ibérica (CGL2011-27343)”. - Synthesys Project (grants FR-TAF-80 and FR-TAF-1735; http://www.synthesys.info/), financed by the European Community Research Infrastructure Action under the FP6 “Structuring the European Research Area” Programme. -

India's Geodynamic Evolution During the Eocene
Article 489 Sunil Bajpai1 and Vivesh V. Kapur2 India’s geodynamic evolution during the Eocene: perspectives on the origin and early evolution of modern mammal orders 1*Department of Earth Sciences, Indian Institute of Technology, Roorkee 247667, India 2 Birbal Sahni Institute of Palaeosciences, Lucknow 226007, India *Corresponding author; Email: [email protected] (Received : 30/01/2019; Revised accepted : 11/09/2019) https://doi.org/10.18814/epiiugs/2020/020031 In recent years, explosion of research in the early and the resulting biotic endemism as well as dispersal during this Tertiary mammals of India has attracted widespread interval. The fossil biota from this interval encompasses the interval from India’s last phase of northward drift until its suturing with Asia. interest because of the importance of this fauna in In recent years it has been shown to represent three distinct understanding biogeographic origins, early evolution, biogeographic domains: Gondwanan, Laurasian, and the endemic and dispersal patterns of several modern mammal component. These components present major biogeographic puzzles orders as well for its paleogeographic implications. and have led to hypotheses that emphasize the fact that the distribution Although Paleocene mammals are yet to be discovered of terrestrial fossil biota provides compelling evidence for the reconstruction of paleogeographic relationships between the in the Indian subcontinent, Indian Early Eocene mammal landmasses (e.g., Ali and Aitchison, 2008; Chatterjee et al., 2017; faunas are now becoming increasingly important in Kapur and Khosla, 2018). debates concerning the origins of several modern The northern (Laurasian) land masses (North America/Europe/ terrestrial orders. In many cases, Eocene mammals China) have been historically considered to be the centre of placental from India represent primitive and stratigraphically evolution because of their rich fossil records from the Paleocene and Eocene. -

The Tempo of Trophic Evolution in Small-Bodied Primates 2 3 4 Running Title: Trophic Evolution in Small-Bodied Primates 5 6 7 Jeremiah E
bioRxiv preprint doi: https://doi.org/10.1101/2020.03.17.996207; this version posted March 20, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-ND 4.0 International license. 1 1 The tempo of trophic evolution in small-bodied primates 2 3 4 Running title: Trophic evolution in small-bodied primates 5 6 7 Jeremiah E. Scott 8 Department of Medical Anatomical Sciences 9 College of Osteopathic Medicine of the Pacific 10 Western University of Health Sciences 11 309 East Second Street 12 Pomona, California 91766 13 U.S.A. 14 [email protected] 15 16 17 Abstract 18 19 Objectives: As a primary trophic strategy, insectivory is uncommon and unevenly 20 distributed across extant primates. This pattern is partly a function of the challenges that 21 insectivory poses for large-bodied primates. In this study, I demonstrate that the uneven 22 distribution is also a consequence of variation in the rate of trophic evolution among small- 23 bodied lineages. 24 Methods: The sample consisted of 307 species classified by primary trophic strategy and 25 body size, creating an ordered three-state character: small-insectivorous, small-herbivorous, 26 and large-herbivorous. I tested for rate heterogeneity by partitioning major clades from the 27 rest of the primate tree and estimating separate rates of transition between herbivory and 28 insectivory for small-bodied lineages in each partition. 29 Results: Bayesian analysis of rate estimates indicates that a model with two rates of trophic 30 evolution provides the best fit to the data. -

Workflow-Simplified Ancestral State Reconstruction of Discrete Traits with Mrbayes in the R Environment
bioRxiv preprint doi: https://doi.org/10.1101/2021.01.10.426107; this version posted January 10, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. MBASR: Workflow-simplified ancestral state reconstruction of discrete traits with MrBayes in the R environment 1,2 Steven Heritage 1 Duke Lemur Center Museum of Natural History, Duke University, Durham, NC, United States 2 Interdepartmental Doctoral Program in Anthropological Science, Stony Brook University, Stony Brook, NY, United States Abstract Ancestral state reconstruction (ASR) is a critical tool in comparative biology. Given a phylogenetic tree and observed trait states for taxa at the tree’s tips, ASR aims to model ancestral trait states at the tree’s internal nodes. Thus, the pattern, tempo, and frequency of a character’s evolution can be considered in a phylogenetic framework. Software programs that perform discrete trait ASR analyses are variably complex in terms of workflow steps, input formats, user interfaces, settings and execution commands, and the organization of output. MBASR (acronym for MrBayes Ancestral States with R) is an R language toolkit that highly automates the ASR workflow and uses the machinery of the popular phylogenetics software MrBayes. Input data files are simple, analysis can be setup and run with a single R function, and statistical output is concisely organized and immediately interpretable. Use requires minimal knowledge of the most basic R commands and no experience with the MrBayes program. -

Vol 31 No14 Final 12-19-07.Indd
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL. 31, NO. 14, PP. 379-385 December 20, 2007 A NEW ADAPOID PRIMATE FROM THE EARLY EOCENE OF INDIA BY KENNETH D. ROSE, RAJENDRA S. RANA, ASHOK SAHNI, AND THIERRY SMITH MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN ANN ARBOR CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY Philip D. Gingerich, Director This series of contributions from the Museum of Paleontology is a medium for publication of papers based chiefl y on collections in the museum. When the number of pages issued is suffi cient to make a volume, a title page plus a table of contents will be sent to libraries on the Museum’s mailing list. This will be sent to individuals on request. A list of separate issues may also be obtained by request. Cor- respondence should be directed to the Publications Secretary, Museum of Paleontology, The University of Michigan, 1109 Geddes Road, Ann Arbor, Michigan 48109-1079 ([email protected]). VOLS. 1-31: Parts of volumes may be obtained if available. Price lists are available upon inquiry. See also: www.paleontology.lsa.umich.edu/Publications/publicationIntro.htm Publications of the Museum of Paleontology are accessible online at: http://deepblue.lib.umich.edu/handle/2027.42/41251 l Text and illustrations ©2007 by the Museum of Paleontology, University of Michigan A NEW ADAPOID PRIMATE FROM THE EARLY EOCENE OF INDIA BY KENNETH D. ROSE1, RAJENDRA S. RANA2, ASHOK SAHNI3, and THIERRY SMITH4 Abstract—A new genus and species of primitive adapoid primate, Asiadapis cambay- ensis, is described based on a dentary from the lower Eocene Cambay Shale ex- posed in the Vastan lignite mine in Gujarat, western India. -

A Fossil Primate of Uncertain Affinities from the Earliest Late Eocene of Egypt
A fossil primate of uncertain affinities from the earliest late Eocene of Egypt Erik R. Seifferta,1, Elwyn L. Simonsb,1, Doug M. Boyerc, Jonathan M. G. Perryd, Timothy M. Ryane, and Hesham M. Sallamf aDepartment of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794-8081; bDivision of Fossil Primates, Duke Lemur Center, Durham, NC 27705; cDepartment of Ecology and Evolution, Stony Brook University, Stony Brook, NY 11794-5245; dDepartment of Anatomy, Midwestern University, Downers Grove, IL 60515; eDepartment of Anthropology, Pennsylvania State University, University Park, PA 16802; and fDepartment of Earth Sciences, University of Oxford, Oxford OX1 3PR, United Kingdom Contributed by Elwyn L. Simons, February 18, 2010 (sent for review January 5, 2010) Paleontological work carried out over the last 3 decades has es- as caenopithecines are likely derived from an independent colo- tablished that three major primate groups were present in the Eocene nization from Europe or Asia. The timing of anthropoids’ arrival in of Africa—anthropoids, adapiforms, and advanced strepsirrhines. Afro-Arabia is unclear, but the presence of multiple anthropoid Here we describe isolated teeth of a previously undocumented pri- species at BQ-2 supports a colonization in the late middle Eocene mate from the earliest late Eocene (≈37 Ma) of northern Egypt, Nos- or earlier. mips aenigmaticus, whose phylogenetic placement within Primates is Here we describe a rare and enigmatic primate species from BQ-2 unclear. Nosmips is smaller than the sympatric adapiform Afradapis that, unlike other primates from theEoceneofAfrica,doesnotfit but is considerably larger than other primate taxa known from the unambiguously into either Anthropoidea or Strepsirrhini on the basis same paleocommunity. -

Paleurafrica Origin of the European Modern Faunas Through Palaeogene Central Africa Collections
PalEurAfrica Origin of the European modern faunas through Palaeogene Central Africa collections BR/121/A3/PalEurAfrica Thierry SMITH (Royal Belgian Institute of Natural Sciences, Belgium) - Thierry DE PUTTER (Royal Museum for Central Africa, Belgium) - Stephen LOUWYE (Ghent University, Ghent, Belgium) - Johan YANS (Namur University, Namur, Belgium) - Nancy STEVENS (Ohio University, Athens, USA) - Annelise FOLIE (Royal Belgian Institute of Natural Sciences, Belgium) Axis 3: Cultural, historical and scientific heritage 11 Project BR/121/A3/PalEurAfrica - Origin of the European modern faunas through Palaeogene Central Africa collections NETWORK PROJECT PALEURAFRICA Origin of the European modern faunas through Palaeogene Central Africa collections Contract - BR/121/A3/PalEurAfrica FINAL REPORT PROMOTORS: Thierry SMITH (Royal Belgian Institute of Natural Sciences, Belgium) Thierry DE PUTTER (Royal Museum for Central Africa, Belgium) Stephen LOUWYE (Ghent University, Ghent, Belgium) Johan YANS (Namur University, Namur, Belgium) Gregg F. GUNNELL† (Duke University, Durham, USA) Nancy STEVENS (Ohio University, Athens, USA) AUTHORS: Thierry SMITH (RBINS), Thierry DE PUTTER (RMCA), Stephen LOUWYE (UGhent), Johan YANS (UNamur), Nancy STEVENS (Ohio Univ.), Annelise FOLIE (RBINS) BRAIN-be (Belgian Research Action through Interdisciplinary Networks) 2 Project BR/121/A3/PalEurAfrica - Origin of the European modern faunas through Palaeogene Central Africa collections Published in 2021 by the Belgian Science Policy Office WTCIII Simon Bolivarlaan 30 Boulevard Simon Bolivar B-1000 Brussels Belgium Tel: +32 (0)2 238 34 11 - Fax: +32 (0)2 230 59 12 http://www.belspo.be http://www.belspo.be/brain-be Contact person: Maaike VANCAUWENBERGHE Tel: +32 (0)2 238 36 78 Neither the Belgian Science Policy Office nor any person acting on behalf of the Belgian Science Policy Office is responsible for the use which might be made of the following information. -

A New Primate from the Late Eocene of Vietnam Illuminates
www.nature.com/scientificreports OPEN A new primate from the late Eocene of Vietnam illuminates unexpected strepsirrhine diversity and evolution in Southeast Asia Olivier Chavasseau1*, Yaowalak Chaimanee1, Stéphane Ducrocq1, Vincent Lazzari1, Phan Dong Pha2,3, Mana Rugbumrung4, Jérôme Surault1, Dang Minh Tuan3 & Jean-Jacques Jaeger1 Sivaladapidae is a poorly known Asian strepsirrhine family originally discovered in Miocene sediments of the Indian subcontinent. Subsequent research has considerably increased the diversity, temporal range, and geographical distribution of this group, now documented from China, Thailand, Myanmar, Pakistan, and India and whose earliest representatives date back to the Middle Eocene. We present here a new taxon of sivaladapid from the Na Duong coal mine in the Latest Middle Eocene-Late Eocene of Vietnam. It represents the frst Eocene primate from Vietnam and the frst medium-sized mammal recovered from this locality, thus documenting a completely new part of the Na Duong paleobiodiversity. This taxon is the largest sivaladapid ever found with an estimated body weight of 5.3 kg and it represents a new subfamily of sivaladapids in exhibiting a very peculiar combination of dental features yet unknown in the fossil record of the family (e.g., retention of four premolars, high-crowned molars with accentuated bunodonty and extreme crest reduction). Besides documenting a complete new part of sivaladapid evolution, its primitive dental formula and derived features shared with the Early Eocene Asiadapidae reinforce the hypothesis of a basal branching of sivaladapids among strepsirrhines. Te basin of Na Duong, in Lang Son province of northern Vietnam, is a pull-apart basin along the Cao Bang-Tien Yen transform fault zone (Fig.