The Stapes of Thrinaxodon Seeley, 1894 and Galesaurus Owen, 1859: a Case of Study for Intraspecific Variability in Basal Cynodonts
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Reptile-Like Physiology in Early Jurassic Stem-Mammals
bioRxiv preprint doi: https://doi.org/10.1101/785360; this version posted October 10, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Title: Reptile-like physiology in Early Jurassic stem-mammals Authors: Elis Newham1*, Pamela G. Gill2,3*, Philippa Brewer3, Michael J. Benton2, Vincent Fernandez4,5, Neil J. Gostling6, David Haberthür7, Jukka Jernvall8, Tuomas Kankanpää9, Aki 5 Kallonen10, Charles Navarro2, Alexandra Pacureanu5, Berit Zeller-Plumhoff11, Kelly Richards12, Kate Robson-Brown13, Philipp Schneider14, Heikki Suhonen10, Paul Tafforeau5, Katherine Williams14, & Ian J. Corfe8*. Affiliations: 10 1School of Physiology, Pharmacology & Neuroscience, University of Bristol, Bristol, UK. 2School of Earth Sciences, University of Bristol, Bristol, UK. 3Earth Science Department, The Natural History Museum, London, UK. 4Core Research Laboratories, The Natural History Museum, London, UK. 5European Synchrotron Radiation Facility, Grenoble, France. 15 6School of Biological Sciences, University of Southampton, Southampton, UK. 7Institute of Anatomy, University of Bern, Bern, Switzerland. 8Institute of Biotechnology, University of Helsinki, Helsinki, Finland. 9Department of Agricultural Sciences, University of Helsinki, Helsinki, Finland. 10Department of Physics, University of Helsinki, Helsinki, Finland. 20 11Helmholtz-Zentrum Geesthacht, Zentrum für Material-und Küstenforschung GmbH Germany. 12Oxford University Museum of Natural History, Oxford, OX1 3PW, UK. 1 bioRxiv preprint doi: https://doi.org/10.1101/785360; this version posted October 10, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 13Department of Anthropology and Archaeology, University of Bristol, Bristol, UK. 14Faculty of Engineering and Physical Sciences, University of Southampton, Southampton, UK. -

Micheli Stefanello
UNIVERSIDADE FEDERAL DE SANTA MARIA CENTRO DE CIÊNCIAS NATURAIS E EXATAS PROGRAMA DE PÓS-GRADUAÇÃO EM BIODIVERSIDADE ANIMAL Micheli Stefanello DESCRIÇÃO E FILOGENIA DE UM NOVO ESPÉCIME DE CINODONTE PROBAINOGNÁTIO DO TRIÁSSICO SUL-BRASILEIRO Santa Maria, RS 2018 Micheli Stefanello DESCRIÇÃO E FILOGENIA DE UM NOVO ESPÉCIME DE CINODONTE PROBAINOGNÁTIO DO TRIÁSSICO SUL-BRASILEIRO Dissertação apresentada ao Curso de Mestrado do Programa de Pós-Graduação em Biodiversidade Animal, Área de Concentração em Sistemática e Biologia Evolutiva, da Universidade Federal de Santa Maria (UFSM, RS), como requisito parcial para obtenção do grau de Mestre em Ciências Biológicas – Área Biodiversidade Animal. Orientador: Prof. Dr. Sérgio Dias da Silva Santa Maria, RS 2018 Micheli Stefanello DESCRIÇÃO E FILOGENIA DE UM NOVO ESPÉCIME DE CINODONTE PROBAINOGNÁTIO DO TRIÁSSICO SUL-BRASILEIRO Dissertação apresentada ao Curso de Mestrado do Programa de Pós-Graduação em Biodiversidade Animal, Área de Concentração em Sistemática e Biologia Evolutiva, da Universidade Federal de Santa Maria (UFSM, RS), como requisito parcial para obtenção do grau de Mestre em Ciências Biológicas – Área Biodiversidade Animal. Aprovada em 28 de fevereiro de 2018: Santa Maria, RS 2018 AGRADECIMENTOS Ao meu orientador, Dr. Sérgio Dias da Silva, pela orientação, por todo o tempo despendido ao longo desse mestrado e por possibilitar meu aprimoramento na área a qual tenho apreço. Aos colegas do Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia da Universidade Federal de Santa Maria (CAPPA/UFSM) e do Laboratório de Paleobiodiversidade Triássica, dessa mesma instituição, pela convivência e por terem ajudado-me de diferentes formas ao longo do mestrado. Em especial, ao colega Rodrigo Temp Müller, pela coleta do espécime (objeto de estudo dessa dissertação), por toda a ajuda com o TNT e com as figuras, e por auxiliar-me de inúmeras formas. -

Studies on Continental Late Triassic Tetrapod Biochronology. I. the Type Locality of Saturnalia Tupiniquim and the Faunal Succession in South Brazil
Journal of South American Earth Sciences 19 (2005) 205–218 www.elsevier.com/locate/jsames Studies on continental Late Triassic tetrapod biochronology. I. The type locality of Saturnalia tupiniquim and the faunal succession in south Brazil Max Cardoso Langer* Departamento de Biologia, FFCLRP, Universidade de Sa˜o Paulo (USP), Av. Bandeirantes 3900, 14040-901 Ribeira˜o Preto, SP, Brazil Received 1 November 2003; accepted 1 January 2005 Abstract Late Triassic deposits of the Parana´ Basin, Rio Grande do Sul, Brazil, encompass a single third-order, tetrapod-bearing sedimentary sequence that includes parts of the Alemoa Member (Santa Maria Formation) and the Caturrita Formation. A rich, diverse succession of terrestrial tetrapod communities is recorded in these sediments, which can be divided into at least three faunal associations. The stem- sauropodomorph Saturnalia tupiniquim was collected in the locality known as ‘Waldsanga’ near the city of Santa Maria. In that area, the deposits of the Alemoa Member yield the ‘Alemoa local fauna,’ which typifies the first association; includes the rhynchosaur Hyperodapedon, aetosaurs, and basal dinosaurs; and is coeval with the lower fauna of the Ischigualasto Formation, Bermejo Basin, NW Argentina. The second association is recorded in deposits of both the Alemoa Member and the Caturrita Formation, characterized by the rhynchosaur ‘Scaphonyx’ sulcognathus and the cynodont Exaeretodon, and correlated with the upper fauna of the Ischigualasto Formation. Various isolated outcrops of the Caturrita Formation yield tetrapod fossils that correspond to post-Ischigualastian faunas but might not belong to a single faunal association. The record of the dicynodont Jachaleria suggests correlations with the lower part of the Los Colorados Formation, NW Argentina, whereas remains of derived tritheledontid cynodonts indicate younger ages. -

Report from the Ina South Africa: SASMT 16 June 2017
Report from the Ina South Africa: SASMT 16 June 2017 Elma Britz The South African Society of Music Teachers (SASMT) was formed in Johannesburg in 1918. The Society can therefore shortly look forward to celebrating its centenary! The SASMT owes its existence to the fact that, since its inception, there was a need to unify the music teaching profession and to further the interest of the profession ‘collectively and individually, besides bringing Music Education and our rich and varied culture to as many people as possible throughout all areas of Southern Africa’ (SASMT handbook revised 2015 - www.sasmt-savmo.org.za). Harry Garvin, a prominent teacher and all-round musician who had come to South Africa from England as a young violinist to lead various light music orchestras and a small group of music teachers felt the need for such a body. The first Annual General Meeting was held in October 1922. The broad outlines of a Constitution were laid down and Garvin was elected president, a position he held for the next nine years. Throughout the years, various membership benefits were established and promulgated. A benevolent fund is available to members who experience unforeseen financial difficulties. A small committee handles deserving applications with the utmost confidentiality. The fund accumulates interest in a special account and a small sum of membership fees is added annually. Many centres collect extra money which is paid into this account. The current editor of our magazine, John Roos, has a small concert venue (Tauromenium) attached to his home where he regularly collects donations from audiences in a newly decorated formerly defunct old cello for this purpose. -

Mammalian Origins Major Groups of Synapsida
Mammalogy 4764 9/16/2009 Chapter 4 “Reptile” Mammalian Origins Early mammal Carnivore Amniota Herbivore Feldhamer Table 4.1 Savage and Long 1986 Major Fig. 3.2, Vaughn, Fig. 4.1, Feldhamer groups Mammalian Origins of Dimetrodon Overview Synapsida Synapsids Pelycosaurs and Therapsids First Mammals Mesozoic Era appear Feldhamer Fig. 4.2 Cenozoic Era radiate Savage and Long 1986 Pelycosaur Skull, jaw musculature, and teeth Cynodontia -- Advanced, predaceous therapsids Pelycosaur Scymnognathus Cynognathus Therapsid Early Cynodont Derived Therapsid/Mammal Primitive Late Cynodont Fig. 3.2, Vaughn Thrinaxodon Fig 4.3 & 4, Feldhamer 1 Mammalogy 4764 9/16/2009 Skeletal transition Extinction of Cynodonts Possibly competition from dinosaurs Pelycosaur Early Cynodonts were dog-size, last surviving were squirrel sized Fig. 4.15 Mammals that survived while Cynodonts went extinct (contemporary) were mouse-sized. Cynodont Thrinaxodon Modern Mammal Fig. 3.5, Vaughn Fig. 4.16c, Early Cynodont Early mammals Changes in land masses Feldhamer 4.11 200 - 250 million years ago Derived characters: Dentary/squamosal jaw articulation Diphyodont dentition 200 MYA 180 MYA Mammary glands Secondary palate Early Mid- Viviparity (loss of eggshell) When? Jurassic Jurassic 65 MYA 135 MYA Early Early Cretaceous Cenozoic Feldhamer 4.5, 4.9 Skull and teeth of mammals 2 Mammalogy 4764 9/16/2009 Teeth and Dentition of Mammals Teeth Heterodont teeth with different functions Differentiated on the basis of function, resulting in increased One of the major keys efficiency acquiring and digesting food. to success of mammals Teeth occur in 3 bones of skull: Teeth of mammals are premaxilla, maxilla, dentary extremely variable with different diets -- more than other taxa Feldhamer et al. -

Xhariep Magisterial District
!. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. XXhhaarriieepp MMaaggiisstteerriiaall DDiissttrriicctt !. !. !. !. !. !. !. !. !. !. TheunissenS ubD istrict !. BARKLY WEST R59 R707 !. ST DEALESVILLE R708 SAPS WINBURG ST R70 !. R370 Lejwelepuitsa SAPS Dealesville R73 Winburg ST ST ST R31 Lejwelepuitsa ST Marquard !. ST LKN12 Boshof !. BRANDFORT Brandfort SAPS !. Soutpan R703 SAPS !. R64 STR64 Magiisteriiall R703 !. ST Sub District Marquard Senekal CAMPBELL ST Kimberley Dealesville ST Ficksburg !. !. !. !. R64 Sub BOSHOF SOUTPAN SAPS KIMBERLEY ST Sub !. !. !. SAPS Diistriict R64 SAPS Brandfort !. SAPS ST R709 Sub District District Sub !. !. Sub ST District Verkeerdevlei MARQUARD Sub District N1 !. SAPS Clocolan !. !. District STR700 KL District -

Gondwana Vertebrate Faunas of India: Their Diversity and Intercontinental Relationships
438 Article 438 by Saswati Bandyopadhyay1* and Sanghamitra Ray2 Gondwana Vertebrate Faunas of India: Their Diversity and Intercontinental Relationships 1Geological Studies Unit, Indian Statistical Institute, 203 B. T. Road, Kolkata 700108, India; email: [email protected] 2Department of Geology and Geophysics, Indian Institute of Technology, Kharagpur 721302, India; email: [email protected] *Corresponding author (Received : 23/12/2018; Revised accepted : 11/09/2019) https://doi.org/10.18814/epiiugs/2020/020028 The twelve Gondwanan stratigraphic horizons of many extant lineages, producing highly diverse terrestrial vertebrates India have yielded varied vertebrate fossils. The oldest in the vacant niches created throughout the world due to the end- Permian extinction event. Diapsids diversified rapidly by the Middle fossil record is the Endothiodon-dominated multitaxic Triassic in to many communities of continental tetrapods, whereas Kundaram fauna, which correlates the Kundaram the non-mammalian synapsids became a minor components for the Formation with several other coeval Late Permian remainder of the Mesozoic Era. The Gondwana basins of peninsular horizons of South Africa, Zambia, Tanzania, India (Fig. 1A) aptly exemplify the diverse vertebrate faunas found Mozambique, Malawi, Madagascar and Brazil. The from the Late Palaeozoic and Mesozoic. During the last few decades much emphasis was given on explorations and excavations of Permian-Triassic transition in India is marked by vertebrate fossils in these basins which have yielded many new fossil distinct taxonomic shift and faunal characteristics and vertebrates, significant both in numbers and diversity of genera, and represented by small-sized holdover fauna of the providing information on their taphonomy, taxonomy, phylogeny, Early Triassic Panchet and Kamthi fauna. -
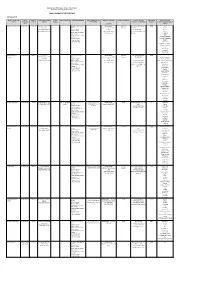
Public Libraries in the Free State
Department of Sport, Arts, Culture & Recreation Directorate Library and Archive Services PUBLIC LIBRARIES IN THE FREE STATE MOTHEO DISTRICT NAME OF FRONTLINE TYPE OF LEVEL OF TOWN/STREET/STREET STAND GPS COORDINATES SERVICES RENDERED SPECIAL SERVICES AND SERVICE STANDARDS POPULATION SERVED CONTACT DETAILS REGISTERED PERIODICALS AND OFFICE FRONTLINE SERVICE NUMBER NUMBER PROGRAMMES CENTER/OFFICE MANAGER MEMBERS NEWSPAPERS AVAILABLE IN OFFICE LIBRARY: (CHARTER) Bainsvlei Public Library Public Library Library Boerneef Street, P O Information and Reference Library hours: 446 142 Ms K Niewoudt Tel: (051) 5525 Car SA Box 37352, Services Ma-Tue, Thu-Fri: 10:00- (Metro) 446-3180 Fair Lady LANGENHOVENPARK, Outreach Services 17:00 Fax: (051) 446-1997 Finesse BLOEMFONTEIN, 9330 Electronic Books Wed: 10:00-18:00 karien.nieuwoudt@mangau Hoezit Government Info Services Sat: 8:30-12:00 ng.co.za Huisgenoot Study Facilities Prescribed books of tertiary Idees Institutions Landbouweekblad Computer Services: National Geographic Internet Access Rapport Word Processing Rooi Rose SA Garden and Home SA Sports Illustrated Sarie The New Age Volksblad Your Family Bloemfontein City Public Library Library c/o 64 Charles Information and Reference Library hours: 443 142 Ms Mpumie Mnyanda 6489 Library Street/West Burger St, P Services Ma-Tue, Thu-Fri: 10:00- (Metro) 051 405 8583 Africa Geographic O Box 1029, Outreach Services 17:00 Architect and Builder BLOEMFONTEIN, 9300 Electronic Books Wed: 10:00-18:00 Tel: (051) 405-8583 Better Homes and Garden n Government Info -

Evolution of the Mammalian Fauces Region and the Origin of Suckling
Evolution of the mammalian fauces region and the origin of suckling The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Crompton, Alfred, Catherine Musinsky, Jose Bonaparte, Bhart- Anjan Bhullar, and Tomasz Owerkowicz. Evolution of the mammalian fauces region and the origin of suckling (2018). Citable link https://nrs.harvard.edu/URN-3:HUL.INSTREPOS:37364482 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA 1 Evolution of the mammalian fauces region and the origin of suckling Alfred Crompton1†, Catherine Musinsky1, Jose Bonaparte2, Bhart-Anjan Bhullar3, Tomasz Owerkowicz4 1: Harvard University, Organismic and Evolutionary Biology Department, Museum of Comparative Zoology, 26 Oxford St, Cambridge, MA 02138 USA 2: Museo Municipal de C. Naturales "C. Ameghino", 6600 MERCEDES – BS. AS. Argentina 3: Department of Geology & Geophysics, Yale University, PO Box 208109, New Haven, CT 06520- 8109 USA 4: Department of Biology, California State University in San Bernadino, 5500 University Pkwy, San Bernardino, CA 92407 USA †corresponding author: ORCID 0000-0001-6008-2587, [email protected], 617-495-3202 Acknowledgments Thanks to Dr. Edgar Allin for his comments on an early draft of this paper. Thanks to the Museum of Comparative Zoology, its Director, Professor James Hanken, and to the Center for Nanoscale Systems at Harvard University for providing the facilities and finances for this research. -

The Role of Fossils in Interpreting the Development of the Karoo Basin
Palaeon!. afr., 33,41-54 (1997) THE ROLE OF FOSSILS IN INTERPRETING THE DEVELOPMENT OF THE KAROO BASIN by P. J. Hancox· & B. S. Rubidge2 IGeology Department, University of the Witwatersrand, Private Bag 3, Wits 2050, South Africa 2Bernard Price Institute for Palaeontological Research, University of the Witwatersrand, Private Bag 3, Wits 2050, South Africa ABSTRACT The Permo-Carboniferous to Jurassic aged rocks oft1:J.e main Karoo Basin ofSouth Africa are world renowned for the wealth of synapsid reptile and early dinosaur fossils, which have allowed a ten-fold biostratigraphic subdivision ofthe Karoo Supergroup to be erected. The role offossils in interpreting the development of the Karoo Basin is not, however, restricted to biostratigraphic studies. Recent integrated sedimentological and palaeontological studies have helped in more precisely defming a number of problematical formational contacts within the Karoo Supergroup, as well as enhancing palaeoenvironmental reconstructions, and basin development models. KEYWORDS: Karoo Basin, Biostratigraphy, Palaeoenvironment, Basin Development. INTRODUCTION Invertebrate remains are important as indicators of The main Karoo Basin of South Africa preserves a facies genesis, including water temperature and salinity, retro-arc foreland basin fill (Cole 1992) deposited in as age indicators, and for their biostratigraphic potential. front of the actively rising Cape Fold Belt (CFB) in Fossil fish are relatively rare in the Karoo Supergroup, southwestern Gondwana. It is the deepest and but where present are useful indicators of gross stratigraphically most complete of several depositories palaeoenvironments (e.g. Keyser 1966) and also have of Permo-Carboniferous to Jurassic age in southern biostratigraphic potential (Jubb 1973; Bender et al. Africa and reflects changing depositional environments 1991). -

A Non-Mammaliaform Cynodont from the Upper Triassic of South Africa: a Therapsid Lazarus Taxon?
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Wits Institutional Repository on DSPACE A non-mammaliaform cynodont from the Upper Triassic of South Africa: a therapsid Lazarus taxon? Fernando Abdala1*, Ross Damiani2, Adam Yates1 & Johann Neveling3 1Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, WITS, 2050 South Africa 2Staatliches Museum für Naturkunde Stuttgart, Rosenstein 1, D-70191, Stuttgart, Germany 3Council for Geoscience, Private Bag X112, Pretoria, 0001 South Africa Received 20 January 2006. Accepted 10 January 2007 The tetrapod record of the ‘Stormberg Group’, including the Lower Elliot Formation, in the South African Karoo is widely dominated by archosaurian reptiles, contrasting with the therapsid dominion of the subjacent Beaufort Group. The only therapsids represented by skeletal remains in the Upper Triassic Lower Elliot Formation are the large traversodontid cynodont Scalenodontoides macrodontes and the recently described tritheledontid cynodont Elliotherium kersteni. Here we present a fragmentary lower jaw that provides evidence of a third type of cynodont for the Upper Triassic of South Africa. The fossil is tentatively assigned to the Diademodontidae. The latter representative of this family is known from the Late Anisian, and its tentative record in the Norian Lower Elliot Formation, if confirmed, will represent a case of Lazarus taxon. Thus, Diademodontidae apparently disappeared from the fossil record by the end of the Anisian and then reappeared in the Norian of South Africa, a stratigraphic interval of some 21 million years. This new cynodont record, together with the recently described Tritheledontidae, show that cynodonts are now the second most diverse tetrapod group in the Lower Elliot fauna. -

Osteohistology of Late Triassic Prozostrodontian Cynodonts from Brazil
Osteohistology of Late Triassic prozostrodontian cynodonts from Brazil Jennifer Botha-Brink1,2, Marina Bento Soares3 and Agustín G. Martinelli3 1 Department of Karoo Palaeontology, National Museum, Bloemfontein, South Africa 2 Department of Zoology and Entomology, University of the Free State, Bloemfontein, South Africa 3 Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil ABSTRACT The Prozostrodontia includes a group of Late Triassic-Early Cretaceous eucynodonts plus the clade Mammaliaformes, in which Mammalia is nested. Analysing their growth patterns is thus important for understanding the evolution of mammalian life histories. Obtaining material for osteohistological analysis is difficult due to the rare and delicate nature of most of the prozostrodontian taxa, much of which comprises mostly of crania or sometimes even only teeth. Here we present a rare opportunity to observe the osteohistology of several postcranial elements of the basal prozostrodontid Prozostrodon brasiliensis, the tritheledontid Irajatherium hernandezi, and the brasilodontids Brasilodon quadrangularis and Brasilitherium riograndensis from the Late Triassic of Brazil (Santa Maria Supersequence). Prozostrodon and Irajatherium reveal similar growth patterns of rapid early growth with annual interruptions later in ontogeny. These interruptions are associated with wide zones of slow growing bone tissue. Brasilodon and Brasilitherium exhibit a mixture of woven-fibered bone tissue and slower growing parallel-fibered and lamellar bone. The slower growing bone tissues are present even during early ontogeny. The relatively slower growth in Brasilodon and Brasilitherium may be related to their small body size compared to Prozostrodon and Irajatherium. These brasilodontids also exhibit osteohistological similarities with the Late Triassic/Early Jurassic mammaliaform Morganucodon and the Late Cretaceous multituberculate mammals Kryptobaatar and Nemegtbaatar.