Supporting Information
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Review of the Diet and Micro-Habitat Values for Wildlife and the Agronomic Potential of Selected Grassland Plant Species
Report Number 697 Review of the diet and micro-habitat values for wildlifeand the agronomic potential of selected grassland plant species English Nature Research Reports working today for nature tomorrow English Nature Research Reports Number 697 Review of the diet and micro-habitat values for wildlife and the agronomic potential of selected grassland plant species S.R. Mortimer, R. Kessock-Philip, S.G. Potts, A.J. Ramsay, S.P.M. Roberts & B.A. Woodcock Centre for Agri-Environmental Research University of Reading, PO Box 237, Earley Gate, Reading RG6 6AR A. Hopkins, A. Gundrey, R. Dunn & J. Tallowin Institute for Grassland and Environmental Research North Wyke Research Station, Okehampton, Devon EX20 2SB J. Vickery & S. Gough British Trust for Ornithology The Nunnery, Thetford, Norfolk IP24 2PU You may reproduce as many additional copies of this report as you like for non-commercial purposes, provided such copies stipulate that copyright remains with English Nature, Northminster House, Peterborough PE1 1UA. However, if you wish to use all or part of this report for commercial purposes, including publishing, you will need to apply for a licence by contacting the Enquiry Service at the above address. Please note this report may also contain third party copyright material. ISSN 0967-876X © Copyright English Nature 2006 Project officer Heather Robertson, Terrestrial Wildlife Team [email protected] Contractor(s) (where appropriate) S.R. Mortimer, R. Kessock-Philip, S.G. Potts, A.J. Ramsay, S.P.M. Roberts & B.A. Woodcock Centre for Agri-Environmental Research, University of Reading, PO Box 237, Earley Gate, Reading RG6 6AR A. -

Iconic Bees: 12 Reports on UK Bee Species
Iconic Bees: 12 reports on UK bee species Bees are vital to the ecology of the UK and provide significant social and economic benefits through crop pollination and maintaining the character of the landscape. Recent years have seen substantial declines in many species of bees within the UK. This report takes a closer look at how 12 ‘iconic’ bee species are faring in each English region, as well as Wales, Northern Ireland and Scotland. Authors Rebecca L. Evans and Simon G. Potts, University of Reading. Photo: © Amelia Collins Contents 1 Summary 2 East England Sea-aster Mining Bee 6 East Midlands Large Garden Bumblebee 10 London Buff-tailed Bumblebee 14 North East Bilberry Bumblebee 18 North West Wall Mason Bee 22 Northern Ireland Northern Colletes 26 Scotland Great Yellow Bumblebee 30 South East England Potter Flower Bee 34 South West England Scabious Bee 38 Wales Large Mason Bee 42 West Midlands Long-horned Bee 46 Yorkshire Tormentil Mining Bee Through collating information on the 12 iconic bee species, common themes have Summary emerged on the causes of decline, and the actions that can be taken to help reverse it. The most pervasive causes of bee species decline are to be found in the way our countryside has changed in the past 60 years. Intensification of grazing regimes, an increase in pesticide use, loss of biodiverse field margins and hedgerows, the trend towards sterile monoculture, insensitive development and the sprawl of towns and cities are the main factors in this. I agree with the need for a comprehensive Bee Action Plan led by the UK Government in order to counteract these causes of decline, as called for by Friends of the Earth. -

Descent of Man
THE DESCENT OF MAN, AND SELECTION IN RELATION TO SEX, BY CHARLES DARWIN, M.A., F.R.S., &c. IN TWO VOLUMES.—VOL. II. WITH ILLUSTRATIONS. LONDON: JOHN MURRAY, ALBEMARLE STREET. 1871. [The right of Translation is reserved.] ERRATA. CONTENTS. PART II. SEXUAL SELECTION-continued. CHAPTER XII. SECONDARY SEXUAL CHARACTERS OF FISHES, AMPHIBIANS, AND REPTILES. FISHES : Courtship and battles of the males — Larger size of the females — Males, bright colours and ornamental appendages; other strange characters — Colours and appendages acquired by the males during the breeding-season alone — Fishes with both sexes brilliantly coloured — Protective colours — The less con- spicuous colours of the female cannot be accounted for on the principle of protection — Male fishes building nests, and taking charge of the ova and young. AMPHIBIANS : Differences in structure and colour between the sexes — Vocal organs. REP- TILES : Chelonians — Crocodiles — Snakes, colours in some cases protective — Lizards, battles of — Ornamental appendages — Strange differences in structure between the sexes — Colours — Sexual differences almost as great as with birds .. Page 1-37 CHAPTER XIII. SECONDARY SEXUAL CHARACTERS OF BIRDS. Sexual differences — Law of battle — Special weapons — Vocal organs—Instrumental music — Love-antics and dances — Deco- rations, permanent and seasonal — Double and single annual moults—Display of ornaments by the males .. .. .. 38-98 vi CONTENTS OF VOL. II. CHAPTER XIV. BIRDS—continued. Choice exerted by the female — Length of courtship — Unpaired birds — Mental qualities and taste for the beautiful — Preference or antipathy shewn by the female for particular males — Vari- ability of birds — Variations sometimes abrupt—Laws of varia- tion — Formation of ocelli — Gradations of character — Case of Peacock, Argus pheasant, and Urosticte . -

Flower Preferences and Pollen Transport Networks for Cavity‐Nesting Solitary Bees: Implications for the Design of Agri‐Envir
Received: 14 February 2018 | Revised: 22 April 2018 | Accepted: 23 April 2018 DOI: 10.1002/ece3.4234 ORIGINAL RESEARCH Flower preferences and pollen transport networks for cavity- nesting solitary bees: Implications for the design of agri- environment schemes Catherine E. A. Gresty1 | Elizabeth Clare2 | Dion S. Devey3 | Robyn S. Cowan3 | Laszlo Csiba3 | Panagiota Malakasi3 | Owen T. Lewis1 | Katherine J. Willis1,3 1Department of Zoology, University of Oxford, Oxford, UK Abstract 2School of Biological and Chemical Floral foraging resources are valuable for pollinator conservation on farmland, and Sciences, Queen Mary University of London, their provision is encouraged by agri- environment schemes in many countries. Across London, UK Europe, wildflower seed mixtures are widely sown on farmland to encourage pollina- 3Royal Botanic Gardens, Kew, Richmond, UK tors, but the extent to which key pollinator groups such as solitary bees exploit and Correspondence benefit from these resources is unclear. We used high- throughput sequencing of 164 Catherine E. A. Gresty, Department of Zoology, New Radcliffe House, Radcliffe pollen samples extracted from the brood cells of six common cavity- nesting solitary Observatory Quarter, 6GG, Woodstock Rd, bee species (Osmia bicornis, Osmia caerulescens, Megachile versicolor, Megachile Oxford OX2, UK. Email: [email protected] ligniseca, Megachile centuncularis and Hylaeus confusus) which are widely distributed across the UK and Europe. We documented their pollen use across 19 farms in south- ern England, UK, revealing their forage plants and examining the structure of their pollen transport networks. Of the 32 plant species included currently in sown wild- flower mixes, 15 were recorded as present within close foraging range of the bees on the study farms, but only Ranunculus acris L. -
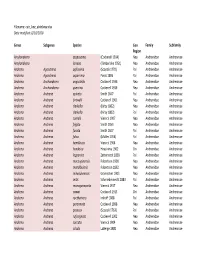
Anthophila List
Filename: cuic_bee_database. -

Hymenoptera, Apoidea)
>lhetian JMfuseum ox4tates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2 2 24 AUGUST I7, I 965 The Biology and Immature Stages of Melitturga clavicornis (Latreille) and of Sphecodes albilabris (Kirby) and the Recognition of the Oxaeidae at the Family Level (Hymenoptera, Apoidea) BYJEROME G. ROZEN, JR.' Michener (1944) divided the andrenid subfamily Panurginae into two tribes, the Panurgini and Melitturgini, with the latter containing the single Old World genus Melitturga. This genus was relegated to tribal status apparently on the grounds that the adults, unlike those of other panurgines, bear certain striking resemblances to the essentially Neo- tropical Oxaeinae of the same family. In 1951 Rozen showed that the male genitalia of Melitturga are unlike those of the Oxaeinae and are not only typical of those of the Panurginae in general but quite like those of the Camptopoeum-Panurgus-Panurginus-Epimethea complex within the sub- family. On the basis of this information, Michener (1954a) abandoned the idea that the genus Melitturga represents a distinct tribe of the Panur- ginae. Recently evidence in the form of the larva of Protoxaea gloriosa Fox (Rozen, 1965) suggested that the Oxaeinae were so unlike other Andreni- dae that they should be removed from the family unless some form inter- 1 Chairman and Associate Curator, Department of Entomology, the American Museum of Natural History. 2 AMERICAN MUSEUM NOVITATES NO. 2224 mediate between the two subfamilies is found. In spite of the structure of the male genitalia, Melitturga is the only known possible intermediary. -

Creating a Pollinator Garden for Native Specialist Bees of New York and the Northeast
Creating a pollinator garden for native specialist bees of New York and the Northeast Maria van Dyke Kristine Boys Rosemarie Parker Robert Wesley Bryan Danforth From Cover Photo: Additional species not readily visible in photo - Baptisia australis, Cornus sp., Heuchera americana, Monarda didyma, Phlox carolina, Solidago nemoralis, Solidago sempervirens, Symphyotrichum pilosum var. pringlii. These shade-loving species are in a nearby bed. Acknowledgements This project was supported by the NYS Natural Heritage Program under the NYS Pollinator Protection Plan and Environmental Protection Fund. In addition, we offer our appreciation to Jarrod Fowler for his research into compiling lists of specialist bees and their host plants in the eastern United States. Creating a Pollinator Garden for Specialist Bees in New York Table of Contents Introduction _________________________________________________________________________ 1 Native bees and plants _________________________________________________________________ 3 Nesting Resources ____________________________________________________________________ 3 Planning your garden __________________________________________________________________ 4 Site assessment and planning: ____________________________________________________ 5 Site preparation: _______________________________________________________________ 5 Design: _______________________________________________________________________ 6 Soil: _________________________________________________________________________ 6 Sun Exposure: _________________________________________________________________ -

Bees and Wasps of the East Sussex South Downs
A SURVEY OF THE BEES AND WASPS OF FIFTEEN CHALK GRASSLAND AND CHALK HEATH SITES WITHIN THE EAST SUSSEX SOUTH DOWNS Steven Falk, 2011 A SURVEY OF THE BEES AND WASPS OF FIFTEEN CHALK GRASSLAND AND CHALK HEATH SITES WITHIN THE EAST SUSSEX SOUTH DOWNS Steven Falk, 2011 Abstract For six years between 2003 and 2008, over 100 site visits were made to fifteen chalk grassland and chalk heath sites within the South Downs of Vice-county 14 (East Sussex). This produced a list of 227 bee and wasp species and revealed the comparative frequency of different species, the comparative richness of different sites and provided a basic insight into how many of the species interact with the South Downs at a site and landscape level. The study revealed that, in addition to the character of the semi-natural grasslands present, the bee and wasp fauna is also influenced by the more intensively-managed agricultural landscapes of the Downs, with many species taking advantage of blossoming hedge shrubs, flowery fallow fields, flowery arable field margins, flowering crops such as Rape, plus plants such as buttercups, thistles and dandelions within relatively improved pasture. Some very rare species were encountered, notably the bee Halictus eurygnathus Blüthgen which had not been seen in Britain since 1946. This was eventually recorded at seven sites and was associated with an abundance of Greater Knapweed. The very rare bees Anthophora retusa (Linnaeus) and Andrena niveata Friese were also observed foraging on several dates during their flight periods, providing a better insight into their ecology and conservation requirements. -

How Habitat Influences Native Pollinators in Intensive Agricultural Landscapes
Small Restoration, Big Impacts: How Habitat Influences Native Pollinators in Intensive Agricultural Landscapes by Caitlin Paterson A Thesis presented to The University of Guelph In partial fulfilment of requirements for the degree of Master of Science in Integrative Biology Guelph, Ontario, Canada © Caitlin Paterson, October, 2014 ABSTRACT SMALL RESTORATION, BIG IMPACTS: HOW HABITAT INFLUENCES NATIVE POLLINATORS IN INTENSIVE AGRICULTURAL LANDSCAPES Caitlin Paterson Advisor: University of Guelph, 2014 Professor A.S. MacDougall This study examined the impact of small-scale restoration of tallgrass prairie on native bee community composition on farms in Southern Ontario, Canada. Three farms with various crops (corn (Zea), Saskatoon berries (Amelanchier alnifolia), squash, pumpkin, zucchini, melon (Curcurbitaceae), soybeans (Glycine), and green beans (Phaseolus)) were surveyed in the summer of 2013. Availability of nesting and foraging habitat for bees was estimated and a combination of pan-trapping and sweep netting was used to capture specimens on 40 plots per farm. Results were analysed using ordination-based techniques in R, and indicated that prairie provides a diverse floral resource on which a complex and abundant array of bees forage. Restored habitat accounted for 33% of the total richness and 72% of the total abundance of native bees. This demonstrates that even small-scale restoration (~10% of each farm) may have an impact on the ability of farms in southern Ontario to support native bees. ACKNOWLEDGMENTS Firstly, thank you to my advisor Andrew MacDougall and the rest of the MacDougall lab for their assistance and guidance with this project for the past two years. I would like to thank the lab/field technicians who assisted me in conducting my field work and identifying and sorting through specimens: Cara Bulger, April Clyburne-Sherin, and Felicia Syer. -

A Catalogue of the Family Andrenidae (Hymenoptera: Apoidea) of Eritrea
Linzer biol. Beitr. 50/1 655-659 27.7.2018 A catalogue of the family Andrenidae (Hymenoptera: Apoidea) of Eritrea Michael MADL A b s t r a c t : In Eritrea the family Andrenidae is represented by two species of the genus Andrena FABRICIUS, 1775 (subfamily Andreninae) and one species each oft he genera Borgatomelissa PATINY, 2000 and Meliturgula FRIESE, 1903 (both subfamily Panurginae). The record of the genus Andrena is doubtful. K e y w o r d s : Andrenidae, Andreninae, Andrena, Panurginae, Borgatomelissa, Meliturgula, Eritrea. Introduction In the Afrotropical region the family Andrenidae is represented by more than 30 species (EARDLEY & URBAN 2010). The Eritrean fauna of of the family Andrenidae is poorly known. The knowledge is based on the material collected by J.K. Lord in 1869 and by P. Magretti in 1900. No further material is known. Up to date two species each of the subfamilies Andreninae (genus Andrena FABRICIUS, 1775) and Panurginae (genera Borgatomelissa PATINY, 2000 and Meliturgula FRIESE, 1903) have been recorded from Eritrea. Abbreviations Afr. reg. .................................. Afrotropical region biogeogr. ......................................... biogeography biol. .......................................................... biology cat. ......................................................... catalogue descr. ................................................... description fig. (figs) ....................................... figure (figures) syn. ......................................................... synonym tab. -

Kew Science Publications for the Academic Year 2017–18
KEW SCIENCE PUBLICATIONS FOR THE ACADEMIC YEAR 2017–18 FOR THE ACADEMIC Kew Science Publications kew.org For the academic year 2017–18 ¥ Z i 9E ' ' . -,i,c-"'.'f'l] Foreword Kew’s mission is to be a global resource in We present these publications under the four plant and fungal knowledge. Kew currently has key questions set out in Kew’s Science Strategy over 300 scientists undertaking collection- 2015–2020: based research and collaborating with more than 400 organisations in over 100 countries What plants and fungi occur to deliver this mission. The knowledge obtained 1 on Earth and how is this from this research is disseminated in a number diversity distributed? p2 of different ways from annual reports (e.g. stateoftheworldsplants.org) and web-based What drivers and processes portals (e.g. plantsoftheworldonline.org) to 2 underpin global plant and academic papers. fungal diversity? p32 In the academic year 2017-2018, Kew scientists, in collaboration with numerous What plant and fungal diversity is national and international research partners, 3 under threat and what needs to be published 358 papers in international peer conserved to provide resilience reviewed journals and books. Here we bring to global change? p54 together the abstracts of some of these papers. Due to space constraints we have Which plants and fungi contribute to included only those which are led by a Kew 4 important ecosystem services, scientist; a full list of publications, however, can sustainable livelihoods and natural be found at kew.org/publications capital and how do we manage them? p72 * Indicates Kew staff or research associate authors. -

Of the Anthophora-Species of Egypt
Prof. Dr. H. PRIESNER A REVIEW OF THE ANTHOPHORA-SPECIES OF EGYPT [Hymenoptera : Apidae] A HEVIEW OF THE ANTHOPHORA-SPECIES OF EGYPT (Hymenoptera : Apidae] by Prof. Dr. H. PRIESNER INTRODUCTION Owing to the difficulties I encountered in trying to identify the Egyptian material of Anthophora in the collections of the Cairo and Ain Shams Universities, I had to penetrate more deeply into this matter, especially when I found that quite a number of species of the local collections were no doubt incorrectly named. With the progress of science, in our particular case with the improvements achieved by finding new distinctive characters and especially in the better relative evaluation of those already known, gained by experience and eye training, there lies upon us the bitter task to criticise and correct our late authorities on this subject who actually did all the spade work that enabled us to start on a considerable higher level of knowledge than that having been at their disposal. Apart from the relatively excellent work of KLUG (1845) and a number of species described by SPINOLA and - much later - GRIBODO, most of the taxonomic work on Anthophora of this country was done by H. FRIESE and A. ALFKEN, their work having been mainly based upon the Apid collections of our late A. ANDRES, while my late friend ALFKEN had also examined specimens he received from the Ministry of Agriculture, Mr. A. ALFIERI and the author. ALFKEN had, obviously with the intention of avoiding the creation of synonyms, identified a number of Egyptian species with such of Asiatic origin.