Xing Et Al. 2018 Goseongornipes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lower Cretaceous Avian-Dominated, Theropod
Lower cretaceous avian-dominated, theropod, thyreophoran, pterosaur and turtle track assemblages from the Tugulu Group, Xinjiang, China: ichnotaxonomy and palaeoecology Lida Xing1,2, Martin G. Lockley3, Chengkai Jia4, Hendrik Klein5, Kecheng Niu6, Lijun Zhang7, Liqi Qi8, Chunyong Chou2, Anthony Romilio9, Donghao Wang2, Yu Zhang2, W Scott Persons10 and Miaoyan Wang2 1 State Key Laboratory of Biogeology and Environmental Geology, China University of Geoscience (Beijing), Beijing, China 2 School of the Earth Sciences and Resources, China University of Geoscience (Beijing), Beijing, China 3 Dinosaur Trackers Research Group, University of Colorado at Denver, Denver, United States 4 Research Institute of Experiment and Detection of Xinjiang Oil Company, PetroChina, Karamay, China 5 Saurierwelt Paläontologisches Museum, Neumarkt, Germany 6 Yingliang Stone Natural History Museum, Nan’an, China 7 Institute of Resources and Environment, Key Laboratory of Biogenic Traces & Sedimentary Minerals of Henan Province, Collaborative Innovation Center of Coalbed Methane and Shale Gas for Central Plains Economic Region, Henan Polytechnic University, Jiaozuo, China 8 Faculty of Petroleum, China University of Petroleum (Beijing) at Karamay, Karamay, China 9 School of Biological Sciences, The University of Queensland, Brisbane, Australia 10 Mace Brown Museum of Natural History, Department of Geology and Environmental Geosciences, College of Charleston, Charleston, United States ABSTRACT Rich tetrapod ichnofaunas, known for more than a decade, from the Huangyangquan Reservoir (Wuerhe District, Karamay City, Xinjiang) have been an abundant source Submitted 10 January 2021 of some of the largest Lower Cretaceous track collections from China. They originate Accepted 26 April 2021 from inland lacustrine clastic exposures of the 581–877 m thick Tugulu Group, 28 May 2021 Published variously divided into four formations and subgroups in the northwestern margin of Corresponding author the Junggar Basin. -
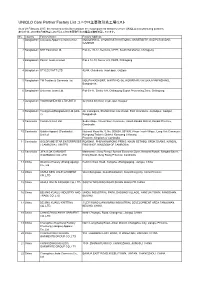
UNIQLO Core Partner Factory List ユニクロ主要取引先工場リスト
UNIQLO Core Partner Factory List ユニクロ主要取引先工場リスト As of 28 February 2017, the factories in this list constitute the major garment factories of core UNIQLO manufacturing partners. 本リストは、2017年2月末時点におけるユニクロ主要取引先の縫製工場を掲載しています。 No. Country Factory Name Factory Address 1 Bangladesh Colossus Apparel Limited unit 2 MOGORKHAL, CHOWRASTA NATIONAL UNIVERSITY, GAZIPUR SADAR, GAZIPUR 2 Bangladesh NHT Fashions Ltd. Plot no. 20-22, Sector-5, CEPZ, South Halishahar, Chittagong 3 Bangladesh Pacific Jeans Limited Plot # 14-19, Sector # 5, CEPZ, Chittagong 4 Bangladesh STYLECRAFT LTD 42/44, Chandona, Joydebpur, Gazipur 5 Bangladesh TM Textiles & Garments Ltd. MOUZA-KASHORE, WARD NO.-06, HOBIRBARI,VALUKA,MYMENSHING, Bangladesh. 6 Bangladesh Universal Jeans Ltd. Plot 09-11, Sector 6/A, Chittagong Export Processing Zone, Chittagong 7 Bangladesh YOUNGONES BD LTD UNIT-II 42 (3rd & 4th floor) Joydevpur, Gazipur 8 Bangladesh Youngones(Bangladesh) Ltd.(Unit- 24, Laxmipura, Shohid chan mia sharak, East Chandona, Joydebpur, Gazipur, 2) Bangladesh 9 Cambodia Cambo Unisoll Ltd. Seda village, Vihear Sour Commune, Ksach Kandal District, Kandal Province, Cambodia 10 Cambodia Golden Apparel (Cambodia) National Road No. 5, No. 005634, 001895, Phsar Trach Village, Long Vek Commune, Limited Kompong Tralarch District, Kompong Chhnang Province, Kingdom of Cambodia. 11 Cambodia GOLDFAME STAR ENTERPRISES ROAD#21, PHUM KAMPONG PRING, KHUM SETHBO, SROK SAANG, KANDAL ( CAMBODIA ) LIMITED PROVINCE, KINGDOM OF CAMBODIA 12 Cambodia JIFA S.OK GARMENT Manhattan ( Svay Rieng ) Special Economic Zone, National Road#, Sangkat Bavet, (CAMBODIA) CO.,LTD Krong Bavet, Svay Rieng Province, Cambodia 13 China Okamoto Hosiery (Zhangjiagang) Renmin West Road, Yangshe, Zhangjiagang, Jiangsu, China Co., Ltd 14 China ANHUI NEW JIALE GARMENT WenChangtown, XuanZhouDistrict, XuanCheng City, Anhui Province CO.,LTD 15 China ANHUI XINLIN FASHION CO.,LTD. -

Ichnotaxonomy of the Eocene Green River Formation
Ichnotaxonomy of the Eocene Green River Formation, Soldier Summit and Spanish Fork Canyon, Uinta Basin, Utah: Interpreting behaviors, lifestyles, and erecting the Cochlichnus Ichnofacies By © 2018 Joshua D. Hogue B.S. Old Dominion University, 2013 Submitted to the graduate degree program in Geology and the Graduate Faculty of the University of Kansas in partial fulfillment of the requirements for the degree of Master of Science. Chair: Dr. Stephen T. Hasiotis Dr. Paul Selden Dr. Georgios Tsoflias Date Defended: May 1, 2018 ii The thesis committee for Joshua D. Hogue certifies that this is the approved version of the following thesis: Ichnotaxonomy of the Eocene Green River Formation, Soldier Summit and Spanish Fork Canyon, Uinta Basin, Utah: Interpreting behaviors, lifestyles, and erecting the Cochlichnus Ichnofacies Chair: Dr. Stephen T. Hasiotis Date Approved: May 1, 2018 iii ABSTRACT The Eocene Green River Formation in the Uinta Basin, Utah, has a diverse ichnofauna. Nineteen ichnogenera and 26 ichnospecies were identified: Acanthichnus cursorius, Alaripeda lofgreni, c.f. Aquatilavipes isp., Aulichnites (A. parkerensis and A. tsouloufeidos isp. nov.), Aviadactyla (c.f. Av. isp. and Av. vialovi), Avipeda phoenix, Cochlichnus (C. anguineus and C. plegmaeidos isp. nov.), Conichnus conichnus, Fuscinapeda texana, Glaciichnium liebegastensis, Glaroseidosichnus ign. nov. gierlowskii isp. nov., Gruipeda (G. fuenzalidae and G. gryponyx), Midorikawapeda ign. nov. semipalmatus isp. nov., Planolites montanus, Presbyorniformipes feduccii, Protovirgularia dichotoma, Sagittichnus linki, Treptichnus (T. bifurcus, T. pedum, and T. vagans), and Tsalavoutichnus ign. nov. (Ts. ericksonii isp. nov. and Ts. leptomonopati isp. nov.). Four ichnocoenoses are represented by the ichnofossils—Cochlichnus, Conichnus, Presbyorniformipes, and Treptichnus—representing dwelling, feeding, grazing, locomotion, predation, pupation, and resting behaviors of organisms in environments at and around the sediment-water-air interface. -

Transmissibility of Hand, Foot, and Mouth Disease in 97 Counties of Jiangsu Province, China, 2015- 2020
Transmissibility of Hand, Foot, and Mouth Disease in 97 Counties of Jiangsu Province, China, 2015- 2020 Wei Zhang Xiamen University Jia Rui Xiamen University Xiaoqing Cheng Jiangsu Provincial Center for Disease Control and Prevention Bin Deng Xiamen University Hesong Zhang Xiamen University Lijing Huang Xiamen University Lexin Zhang Xiamen University Simiao Zuo Xiamen University Junru Li Xiamen University XingCheng Huang Xiamen University Yanhua Su Xiamen University Benhua Zhao Xiamen University Yan Niu Chinese Center for Disease Control and Prevention, Beijing City, People’s Republic of China Hongwei Li Xiamen University Jian-li Hu Jiangsu Provincial Center for Disease Control and Prevention Tianmu Chen ( [email protected] ) Page 1/30 Xiamen University Research Article Keywords: Hand foot mouth disease, Jiangsu Province, model, transmissibility, effective reproduction number Posted Date: July 30th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-752604/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 2/30 Abstract Background: Hand, foot, and mouth disease (HFMD) has been a serious disease burden in the Asia Pacic region represented by China, and the transmission characteristics of HFMD in regions haven’t been clear. This study calculated the transmissibility of HFMD at county levels in Jiangsu Province, China, analyzed the differences of transmissibility and explored the reasons. Methods: We built susceptible-exposed-infectious-asymptomatic-removed (SEIAR) model for seasonal characteristics of HFMD, estimated effective reproduction number (Reff) by tting the incidence of HFMD in 97 counties of Jiangsu Province from 2015 to 2020, compared incidence rate and transmissibility in different counties by non -parametric test, rapid cluster analysis and rank-sum ratio. -

Seroepidemiology of Human Enterovirus71 and Coxsackievirusa16 in Jiangsu Province, China
Ji et al. Virology Journal 2012, 9:248 http://www.virologyj.com/content/9/1/248 RESEARCH Open Access Seroepidemiology of human enterovirus71 and coxsackievirusA16 in Jiangsu province, China Hong Ji1, Liang Li1, YanMing Liu2, HengMing Ge3, XuShan Wang4, JianLi Hu1, Bin Wu1, JianGuang Fu1, ZhenYu Zhang3, XiaoQin Chen3, MingLei Zhang3, Qiang Ding4, WenBo Xu2, FenYang Tang1, MingHao Zhou1, Hua Wang1 and FengCai Zhu1* Abstract Background: The major etiology of hand, foot and mouth disease (HFMD) is infection with human enterovirus A (HEV-A). Among subtypes of HEV-A, coxsackievirusA16 (CoxA16) and enterovirus 71 (EV71) are major causes for recurrent HFMD among infants and children in Jiangsu Province, mainland China. Here, we analyzed maternal antibodies between prenatal women and their neonates, to determine age-specific seroprevalence of human EV71 and CoxA16 infections in infants and children aged 0 to 15 years. The results may facilitate the development of immunization against HFMD. Methods: This study used cross-section of 40 pairs of pregnant women and neonates and 800 subjects aged 1 month to 15 years old. Micro-dose cytopathogenic effects measured neutralizing antibodies against EV71 and CoxA16. Chi-square test compared seroprevalence rates between age groups and McNemar test, paired-Samples t-test and independent-samples t-test analyzed differences of geometric mean titers. Results: A strong correlation between titers of neutralizing antibody against EV71 and CoxA16 in prenatal women and neonates was observed (rEV71 = 0.67, rCoxA16 = 0.56, respectively, p < 0.05). Seroprevalence rates of anti-EV71 antibody gradually decreased with age between 0 to 6 months old, remained low between 7 to 11 months (5.0–10.0%), and increased between 1 and 4 years (22.5–87.5%). -
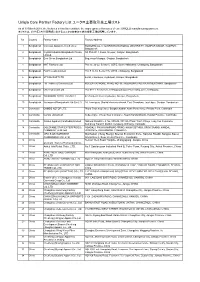
Uniqlo Core Partner Factory List ユニクロ主要取引先工場リスト
Uniqlo Core Partner Factory List ユニクロ主要取引先工場リスト As of 30 March 2018, the factories in this list constitute the major garment factories of core UNIQLO manufacturing partners. 本リストは、2018年3月30日時点におけるユニクロ主要取引先の縫製工場を掲載しています。 No. Country Factory Name Factory Address 1 Bangladesh Colossus Apparel Limited unit 2 MOGORKHAL, CHOWRASTA NATIONAL UNIVERSITY, GAZIPUR SADAR, GAZIPUR, Bangladesh 2 Bangladesh Crystal Industrial Bangladesh Private SA Plot-2013, Kewa, Sreepur, Gazipur, Bangladesh Limited. 3 Bangladesh Ever Smart Bangladesh Ltd. Begumour Mirzapur, Gazipur, Bangladesh 4 Bangladesh NHT Fashions Ltd. Plot no. 20-22, Sector-5, CEPZ, South Halishahar, Chittagong, Bangladesh 5 Bangladesh Pacific Jeans Limited Plot # 14-19, Sector # 5, CEPZ, Chittagong, Bangladesh 6 Bangladesh STYLECRAFT LTD 42/44, Chandona, Joydebpur, Gazipur, Bangladesh 7 Bangladesh TM Textiles & Garments Ltd. MOUZA-KASHORE, WARD NO.-06, HOBIRBARI,VALUKA,MYMENSHING, Bangladesh 8 Bangladesh Universal Jeans Ltd. Plot 09-11, Sector 6/A, Chittagong Export Processing Zone, Chittagong 9 Bangladesh YOUNGONES BD LTD UNIT-II 42 (3rd & 4th floor) Joydevpur, Gazipur, Bangladesh 10 Bangladesh Youngones(Bangladesh) Ltd.(Unit-2) 24, Laxmipura, Shohid chan mia sharak, East Chandona, Joydebpur, Gazipur, Bangladesh 11 Cambodia CAMBO KOTOP LTD. Phum Trapeang Chrey, Sangkat Kakab, Khan Porsenchey, Phnom Penh, Cambodia. 12 Cambodia Cambo Unisoll Ltd. Seda village, Vihear Sour Commune, Ksach Kandal District, Kandal Province, Cambodia 13 Cambodia Golden Apparel (Cambodia) Limited National Road No. -

Cretaceous Research 74 (2017) 155E164
Cretaceous Research 74 (2017) 155e164 Contents lists available at ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/CretRes Mid-Cretaceous dinosaur track assemblage from the Tongfosi Formation of China: Comparison with the track assemblage of South Korea * Lida Xing a, b, , Martin G. Lockley c, Kyung Soo Kim d, Hendrik Klein e, Masaki Matsukawa f, Jong Deock Lim g, W. Scott Persons IV h, Xing Xu i a State Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences, Beijing 100083, China b School of the Earth Sciences and Resources, China University of Geosciences, Beijing 100083, China c Dinosaur Trackers Research Group, University of Colorado Denver, PO Box 173364, Denver, CO 80217, USA d Department of Science Education, Chinju National University of Education, Shinan-dong, Jinju, Kyungnam 660-756, South Korea e Saurierwelt Palaontologisches€ Museum, Alte Richt 7, D-92318 Neumarkt, Germany f Department of Environmental Sciences, Tokyo Gakugei University, Koganei, Tokyo 184-8501, Japan g Natural Heritage Center, National Research Institute of Cultural Heritage, 927 Yudeng-ro, Seo-gu, Daejon 302-834, South Korea h Department of Biological Sciences, University of Alberta, 11455 Saskatchewan Drive, Edmonton, Alberta T6G 2E9, Canada i Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China article info abstract Article history: A Yanji Basin tracksite in the Cretaceous Tongfosi Formation in the Yanbian Korean Autonomous Pre- Received 14 October 2016 fecture, Jilin Province was reinvestigated twenty years after the original study. Received in revised form The re-examination confirms the presence of tracks tentatively referred to ornithopods and theropod 15 February 2017 tracks including several not previously reported. -

Bbm:978-94-017-9597-5/1.Pdf
Index A Arenicolites isp . , 21 , 46 , 64 , 70 , 85 , 88 Abarenicola pacifi ca , 105 Aristopentapodiscus , 373 Acanthichnus , 454 Artichnus , 21 , 43 , 93 Acropentapodiscus , 373 Artichnus pholeoides , 9 4 Acunaichnus dorregoensis , 390 Asaphoidichnus , 454 Ademosynidae , 195 Asencio Formation , 344 Adephagan beetles , 201 Asiatoceratodus , 238 Agua de la Peña Group , 192 Asteriacites , 9 , 21 , 43 , 44 , 66 Alcyonidiopsis , 280 , 283 Asteriacites lumbricalis , 42 , 67 Allocotichnus , 454 Asterosoma , 21 , 46 , 47 , 52 , 63 , 65 , 68 , 69 , 76 , Ameghinichnus , 372 , 373 78 , 85 , 89 , 95 Ameghinichnus patagonicu s , 372 Asterosoma coxii , 5 2 Amphibiopodiscus , 373 Asterosoma ludwigae , 5 2 Anachalcos mfwangani , 346 Asterosoma radiciformis , 5 2 Anchisauripus , 6 , 8 Asterosoma striata , 5 2 Ancorichnus , 4 5 Asterosoma surlyki , 5 2 Angulata Zone , 7 , 11 Atreipus , 6 , 8 , 147 Angulichnus , 454 Australopithecus , 413 Anomoepus , 6–8 Australopiths , 411 Anthropoidipes ameriborealis , 437 Avolatichnium , 192 Anyao Formation , 198 Azolla ferns , 202 Apatopus , 5 , 6 , 146 Arachnomorphichnus , 454 Arachnostega , 21 , 45 , 46 , 49 , 61 B Aragonitermes teruelensis , 335 Baissatermes lapideus , 335 Arariperhinus monnei , 335 Baissoferidae , 211 , 216 Archaeonassa , 34 , 62 , 223 , 454 Balanoglossites , 4 6 Archaeonassa fossulata , 192 Barberichnus bonaerensis , 304 Archeoentomichnus metapolypholeos , 320 Barrancapus , 5 Archosaur trackways , 6 Bathichnus paramoudrae , 8 0 Archosaurs , 137 , 138 , 140 , 143 , 145–147 , 160 Batrachopus , 6 -

Spatial and Temporal Evaluation of Ecological Footprint Intensity of Jiangsu Province at the County-Level Scale
International Journal of Environmental Research and Public Health Article Spatial and Temporal Evaluation of Ecological Footprint Intensity of Jiangsu Province at the County-Level Scale Decun Wu 1 and Jinping Liu 2,* 1 School of Public Administration and Sociology, Jiangsu Normal University, Xuzhou 221116, China; [email protected] 2 School of Economics and Management, China University of Mining and Technology, Xuzhou 221116, China * Correspondence: [email protected] Received: 10 September 2020; Accepted: 23 October 2020; Published: 26 October 2020 Abstract: Due to the high ecological pressure that exists in the process of rapid economic development in Jiangsu Province, it is necessary to evaluate its ecological footprint intensity (EFI). This article focuses on ecological footprint intensity analysis at the county scale. We used county-level data to evaluate the spatial distributions and temporal trends of the ecological footprint intensity in Jiangsu’s counties from 1995 to 2015. The temporal trends of counties are divided into five types: linear declining type, N-shape type, inverted-N type, U-shape type and inverted-U shape type. It was discovered that the proportions of the carbon footprint intensity were maintained or increased in most counties. Exploratory spatial data analysis shows that there was a certain regularity of the EFI spatial distributions, i.e., a gradient decrease from north to south, and there was a decline in the spatial heterogeneity of EFI in Jiangsu’s counties over time. The global Moran’s index (Moran’s I) and local spatial association index (LISA) are used to analyze both the global and local spatial correlation of EFIs among counties of Jiangsu Province. -

Interpreting Behavior from Early Cretaceous Bird Tracks and the Morphology of Bird Feet and Trackways
INTERPRETING BEHAVIOR FROM EARLY CRETACEOUS BIRD TRACKS AND THE MORPHOLOGY OF BIRD FEET AND TRACKWAYS By ©2009 Amanda Renee Falk B.S., Lake Superior State University, 2007 Submitted to the Department of Geology and the Faculty of the Graduate School of the University of Kansas In partial fulfillment of the requirements for the degree of Master of Science Advisory Committee: ______________________________ Co-Chairman: Stephen T. Hasiotis ______________________________ Co-Chairman: Larry D. Martin ______________________________ J. F. Devlin Date Defended: September 15th, 2009 The thesis committee for Amanda R. Falk certifies that this is the approved version of the following thesis: INTERPRETING BEHAVIOR FROM EARLY CRETACEOUS BIRD TRACKS AND THE MORPHOLOGY OF BIRD FEET AND TRACKWAYS Advisory Committee: ____________________________ Stephen T. Hasiotis, Chairman ____________________________ Larry D. Martin, Co-Chairman ____________________________ J. F. Devlin Date approved: _ September 15th, 2009_ ii ABSTRACT Amanda R. Falk Department of Geology, September 2009 University of Kansas Bird tracks were studied from the Lower Cretaceous Lakota Formation in South Dakota, USA, and the Lower Cretaceous Haman Formation, South Korea. Behaviors documented from the Lakota Formation included: (1) a takeoff behavior represented by a trackway terminating in two subparallel tracks; (2) circular walking; and (3) the courtship display high stepping. Behaviors documented from the Haman Formation included: (1) a low-angle landing in which the hallux toe was dragged; (2) pecking and probing behaviors; and (3) flapping-assisted hopping during walking. The invertebrate trace fossil Cochlichnus was associated the avian tracks from the Lakota Formation. No traces of pecking or probing were associated with Cochlichnus. The invertebrate trace fossils Cochlichnus, Arenicholites, and Steinichnus were found associated the bird tracks from the Haman Formation. -

Multiple Parallel Deinonychosaurian Trackways from a Diverse Dinosaur Track Assemblage of the Lower Cretaceous Dasheng Group of Shandong Province, China
Accepted Manuscript Multiple parallel deinonychosaurian trackways from a diverse dinosaur track assemblage of the Lower Cretaceous Dasheng Group of Shandong Province, China Lida Xing, Martin G. Lockley, Ying Guo, Hendrik Klein, Junqiang Zhang, Li Zhang, W. Scott Persons, IV, Anthony Romilio, Yonggang Tang, Xiaoli Wang PII: S0195-6671(17)30544-X DOI: 10.1016/j.cretres.2018.04.005 Reference: YCRES 3854 To appear in: Cretaceous Research Received Date: 16 December 2017 Revised Date: 1 April 2018 Accepted Date: 5 April 2018 Please cite this article as: Xing, L., Lockley, M.G., Guo, Y., Klein, H., Zhang, J., Zhang, L., Persons IV., , W.S., Romilio, A., Tang, Y., Wang, X., Multiple parallel deinonychosaurian trackways from a diverse dinosaur track assemblage of the Lower Cretaceous Dasheng Group of Shandong Province, China, Cretaceous Research (2018), doi: 10.1016/j.cretres.2018.04.005. This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. ACCEPTED MANUSCRIPT Multiple parallel deinonychosaurian trackways from a diverse dinosaur track assemblage of the Lower Cretaceous Dasheng Group of Shandong Province, China Lida Xing a b c , Martin G. Lockley d, Ying Guo e, Hendrik Klein f, Junqiang Zhang e, Li Zhang g, W. -

Subsidy Programs
SUBSIDIES REQUEST FROM THE UNITED STATES TO CHINA PURSUANT TO ARTICLE 25.10 OF THE AGREEMENT The following communication, dated 15 April 2016, is being circulated at the request of the Delegation of the United States. _______________ In the report that it prepared for China's most recent Trade Policy Review, held in July 2014, the Secretariat included information that it had uncovered on 30 support programs for China's fisheries sector.1 The Secretariat's Report noted that China had not notified any of these support programs to the Committee on Subsidies and Countervailing Measures (Committee) and that China could not verify any of the information provided to it by the Secretariat. During the ensuing Trade Policy Review meetings, China responded to Members' questions about these support programs by stating that it needed more time to identify and verify information regarding these support programs. In April of last year, the United States submitted questions to China with respect to these programs and other fishery support measures the United States had uncovered through its own efforts.2 Once again, despite its obligation under Article 25.9 of the Agreement to provide answers "as quickly as possible and in a comprehensive manner", China to date has not meaningfully responded to the United States' request and has refused repeated requests to meet bilaterally to discuss the issue, and the issue of subsidy notifications more generally. In November of last year, China submitted its latest subsidy notification covering the period 2009 through 2014.3 In this notification, China did not include: (1) any of the fishery subsidy programs identified in China's 2014 TPR report, (2) any of the additional measures identified by the United States in its Article 25.8 submission or (3) any of the fisheries subsidies measures identified in the 2014 Article 25.10 submission of the United States4.