Ecology, Survey and Management of Forest Insects1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Forestry Department Food and Agriculture Organization of the United Nations
Forestry Department Food and Agriculture Organization of the United Nations Forest Health & Biosecurity Working Papers OVERVIEW OF FOREST PESTS ROMANIA January 2007 Forest Resources Development Service Working Paper FBS/28E Forest Management Division FAO, Rome, Italy Forestry Department DISCLAIMER The aim of this document is to give an overview of the forest pest1 situation in Romania. It is not intended to be a comprehensive review. The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. © FAO 2007 1 Pest: Any species, strain or biotype of plant, animal or pathogenic agent injurious to plants or plant products (FAO, 2004). Overview of forest pests - Romania TABLE OF CONTENTS Introduction..................................................................................................................... 1 Forest pests and diseases................................................................................................. 1 Naturally regenerating forests..................................................................................... 1 Insects ..................................................................................................................... 1 Diseases................................................................................................................ -

Succession of Coleoptera on Freshly Killed
Louisiana State University LSU Digital Commons LSU Master's Theses Graduate School 2008 Succession of Coleoptera on freshly killed loblolly pine (Pinus taeda L.) and southern red oak (Quercus falcata Michaux) in Louisiana Stephanie Gil Louisiana State University and Agricultural and Mechanical College, [email protected] Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_theses Part of the Entomology Commons Recommended Citation Gil, Stephanie, "Succession of Coleoptera on freshly killed loblolly pine (Pinus taeda L.) and southern red oak (Quercus falcata Michaux) in Louisiana" (2008). LSU Master's Theses. 1067. https://digitalcommons.lsu.edu/gradschool_theses/1067 This Thesis is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Master's Theses by an authorized graduate school editor of LSU Digital Commons. For more information, please contact [email protected]. SUCCESSIO OF COLEOPTERA O FRESHLY KILLED LOBLOLLY PIE (PIUS TAEDA L.) AD SOUTHER RED OAK ( QUERCUS FALCATA MICHAUX) I LOUISIAA A Thesis Submitted to the Graduate Faculty of the Louisiana State University and Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of Master of Science in The Department of Entomology by Stephanie Gil B. S. University of New Orleans, 2002 B. A. University of New Orleans, 2002 May 2008 DEDICATIO This thesis is dedicated to my parents who have sacrificed all to give me and my siblings a proper education. I am indebted to my entire family for the moral support and prayers throughout my years of education. My mother and Aunt Gloria will have several extra free hours a week now that I am graduating. -

Local and Regional Influences on Arthropod Community
LOCAL AND REGIONAL INFLUENCES ON ARTHROPOD COMMUNITY STRUCTURE AND SPECIES COMPOSITION ON METROSIDEROS POLYMORPHA IN THE HAWAIIAN ISLANDS A DISSERTATION SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAI'I IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN ZOOLOGY (ECOLOGY, EVOLUTION AND CONSERVATION BIOLOGy) AUGUST 2004 By Daniel S. Gruner Dissertation Committee: Andrew D. Taylor, Chairperson John J. Ewel David Foote Leonard H. Freed Robert A. Kinzie Daniel Blaine © Copyright 2004 by Daniel Stephen Gruner All Rights Reserved. 111 DEDICATION This dissertation is dedicated to all the Hawaiian arthropods who gave their lives for the advancement ofscience and conservation. IV ACKNOWLEDGEMENTS Fellowship support was provided through the Science to Achieve Results program of the U.S. Environmental Protection Agency, and training grants from the John D. and Catherine T. MacArthur Foundation and the National Science Foundation (DGE-9355055 & DUE-9979656) to the Ecology, Evolution and Conservation Biology (EECB) Program of the University of Hawai'i at Manoa. I was also supported by research assistantships through the U.S. Department of Agriculture (A.D. Taylor) and the Water Resources Research Center (RA. Kay). I am grateful for scholarships from the Watson T. Yoshimoto Foundation and the ARCS Foundation, and research grants from the EECB Program, Sigma Xi, the Hawai'i Audubon Society, the David and Lucille Packard Foundation (through the Secretariat for Conservation Biology), and the NSF Doctoral Dissertation Improvement Grant program (DEB-0073055). The Environmental Leadership Program provided important training, funds, and community, and I am fortunate to be involved with this network. -

T1)E Bedford,1)Ire Naturaii,T 45
T1)e Bedford,1)ire NaturaIi,t 45 Journal for the year 1990 Bedfordshire Natural History Society 1991 'ISSN 0951 8959 I BEDFORDSHffiE NATURAL HISTORY SOCIETY 1991 Chairman: Mr D. Anderson, 88 Eastmoor Park, Harpenden, Herts ALS 1BP Honorary Secretary: Mr M.C. Williams, 2 Ive! Close, Barton-le-Clay, Bedford MK4S 4NT Honorary Treasurer: MrJ.D. Burchmore, 91 Sundon Road, Harlington, Dunstable, Beds LUS 6LW Honorary Editor (Bedfordshire Naturalist): Mr C.R. Boon, 7 Duck End Lane, Maulden, Bedford MK4S 2DL Honorary Membership Secretary: Mrs M.]. Sheridan, 28 Chestnut Hill, Linslade, Leighton Buzzard, Beds LU7 7TR Honorary Scientific Committee Secretary: Miss R.A. Brind, 46 Mallard Hill, Bedford MK41 7QS Council (in addition to the above): Dr A. Aldhous MrS. Cham DrP. Hyman DrD. Allen MsJ. Childs Dr P. Madgett MrC. Baker Mr W. Drayton MrP. Soper Honorary Editor (Muntjac): Ms C. Aldridge, 9 Cowper Court, Markyate, Herts AL3 8HR Committees appointed by Council: Finance: Mr]. Burchmore (Sec.), MrD. Anderson, Miss R. Brind, Mrs M. Sheridan, Mr P. Wilkinson, Mr M. Williams. Scientific: Miss R. Brind (Sec.), Mr C. Boon, Dr G. Bellamy, Mr S. Cham, Miss A. Day, DrP. Hyman, MrJ. Knowles, MrD. Kramer, DrB. Nau, MrE. Newman, Mr A. Outen, MrP. Trodd. Development: Mrs A. Adams (Sec.), MrJ. Adams (Chairman), Ms C. Aldridge (Deputy Chairman), Mrs B. Chandler, Mr M. Chandler, Ms]. Childs, Mr A. Dickens, MrsJ. Dickens, Mr P. Soper. Programme: MrJ. Adams, Mr C. Baker, MrD. Green, MrD. Rands, Mrs M. Sheridan. Trustees (appointed under Rule 13): Mr M. Chandler, Mr D. Green, Mrs B. -

The Curculionoidea of the Maltese Islands (Central Mediterranean) (Coleoptera)
BULLETIN OF THE ENTOMOLOGICAL SOCIETY OF MALTA (2010) Vol. 3 : 55-143 The Curculionoidea of the Maltese Islands (Central Mediterranean) (Coleoptera) David MIFSUD1 & Enzo COLONNELLI2 ABSTRACT. The Curculionoidea of the families Anthribidae, Rhynchitidae, Apionidae, Nanophyidae, Brachyceridae, Curculionidae, Erirhinidae, Raymondionymidae, Dryophthoridae and Scolytidae from the Maltese islands are reviewed. A total of 182 species are included, of which the following 51 species represent new records for this archipelago: Araecerus fasciculatus and Noxius curtirostris in Anthribidae; Protapion interjectum and Taeniapion rufulum in Apionidae; Corimalia centromaculata and C. tamarisci in Nanophyidae; Amaurorhinus bewickianus, A. sp. nr. paganettii, Brachypera fallax, B. lunata, B. zoilus, Ceutorhynchus leprieuri, Charagmus gressorius, Coniatus tamarisci, Coniocleonus pseudobliquus, Conorhynchus brevirostris, Cosmobaris alboseriata, C. scolopacea, Derelomus chamaeropis, Echinodera sp. nr. variegata, Hypera sp. nr. tenuirostris, Hypurus bertrandi, Larinus scolymi, Leptolepurus meridionalis, Limobius mixtus, Lixus brevirostris, L. punctiventris, L. vilis, Naupactus cervinus, Otiorhynchus armatus, O. liguricus, Rhamphus oxyacanthae, Rhinusa antirrhini, R. herbarum, R. moroderi, Sharpia rubida, Sibinia femoralis, Smicronyx albosquamosus, S. brevicornis, S. rufipennis, Stenocarus ruficornis, Styphloderes exsculptus, Trichosirocalus centrimacula, Tychius argentatus, T. bicolor, T. pauperculus and T. pusillus in Curculionidae; Sitophilus zeamais and -

Layman's Report
THE RED LIST STATUS OF EUROPE’S OVERLOOKED SPECIES LIFE14 PRE BE 001 – Layman’s report This project was co-funded by the European Union under the LIFE Financial Instrument and the Grant Agreement n. LIFE14 PRE BE 001 About IUCN Created in 1948, IUCN represents one of the world’s largest and most diverse environmental networks. It harnesses the experience, resources and reach of more than 1,300 member organisations and the input of over 15,000 volunteer experts, organised in six commissions. IUCN is the global authority on the status of the natural world and the measures needed to safeguard it. The IUCN Global Species Programme supports the activities of the IUCN Species Survival Commission and individual Specialist Groups, as well as implementing global species conservation initiatives. It is an integral part of the IUCN Secretariat and is managed from IUCN’s international headquarters in Gland, Switzerland. What is the IUCN Red List? The European Red List The IUCN Red List of Threatened SpeciesTM is the When conducting regional or national assessments, world’s most comprehensive information source on and to ensure that the criteria are applied the extinction risk of plant and animal species. It is a appropriately at such scales, the IUCN has compilation of the conservation status of species at developed the Guidelines for Application of IUCN Red the global level, based on the best scientific List Criteria at Regional Levels.1 information available. The IUCN Red List Categories and Criteria are based on a set of quantitative criteria linked to population trends, size and structure, threats, and geographic ranges of species. -

Disturbance and Recovery of Litter Fauna: a Contribution to Environmental Conservation
Disturbance and recovery of litter fauna: a contribution to environmental conservation Vincent Comor Disturbance and recovery of litter fauna: a contribution to environmental conservation Vincent Comor Thesis committee PhD promotors Prof. dr. Herbert H.T. Prins Professor of Resource Ecology Wageningen University Prof. dr. Steven de Bie Professor of Sustainable Use of Living Resources Wageningen University PhD supervisor Dr. Frank van Langevelde Assistant Professor, Resource Ecology Group Wageningen University Other members Prof. dr. Lijbert Brussaard, Wageningen University Prof. dr. Peter C. de Ruiter, Wageningen University Prof. dr. Nico M. van Straalen, Vrije Universiteit, Amsterdam Prof. dr. Wim H. van der Putten, Nederlands Instituut voor Ecologie, Wageningen This research was conducted under the auspices of the C.T. de Wit Graduate School of Production Ecology & Resource Conservation Disturbance and recovery of litter fauna: a contribution to environmental conservation Vincent Comor Thesis submitted in fulfilment of the requirements for the degree of doctor at Wageningen University by the authority of the Rector Magnificus Prof. dr. M.J. Kropff, in the presence of the Thesis Committee appointed by the Academic Board to be defended in public on Monday 21 October 2013 at 11 a.m. in the Aula Vincent Comor Disturbance and recovery of litter fauna: a contribution to environmental conservation 114 pages Thesis, Wageningen University, Wageningen, The Netherlands (2013) With references, with summaries in English and Dutch ISBN 978-94-6173-749-6 Propositions 1. The environmental filters created by constraining environmental conditions may influence a species assembly to be driven by deterministic processes rather than stochastic ones. (this thesis) 2. High species richness promotes the resistance of communities to disturbance, but high species abundance does not. -

Coleoptera: Introduction and Key to Families
Royal Entomological Society HANDBOOKS FOR THE IDENTIFICATION OF BRITISH INSECTS To purchase current handbooks and to download out-of-print parts visit: http://www.royensoc.co.uk/publications/index.htm This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 2.0 UK: England & Wales License. Copyright © Royal Entomological Society 2012 ROYAL ENTOMOLOGICAL SOCIETY OF LONDON Vol. IV. Part 1. HANDBOOKS FOR THE IDENTIFICATION OF BRITISH INSECTS COLEOPTERA INTRODUCTION AND KEYS TO FAMILIES By R. A. CROWSON LONDON Published by the Society and Sold at its Rooms 41, Queen's Gate, S.W. 7 31st December, 1956 Price-res. c~ . HANDBOOKS FOR THE IDENTIFICATION OF BRITISH INSECTS The aim of this series of publications is to provide illustrated keys to the whole of the British Insects (in so far as this is possible), in ten volumes, as follows : I. Part 1. General Introduction. Part 9. Ephemeroptera. , 2. Thysanura. 10. Odonata. , 3. Protura. , 11. Thysanoptera. 4. Collembola. , 12. Neuroptera. , 5. Dermaptera and , 13. Mecoptera. Orthoptera. , 14. Trichoptera. , 6. Plecoptera. , 15. Strepsiptera. , 7. Psocoptera. , 16. Siphonaptera. , 8. Anoplura. 11. Hemiptera. Ill. Lepidoptera. IV. and V. Coleoptera. VI. Hymenoptera : Symphyta and Aculeata. VII. Hymenoptera: Ichneumonoidea. VIII. Hymenoptera : Cynipoidea, Chalcidoidea, and Serphoidea. IX. Diptera: Nematocera and Brachycera. X. Diptera: Cyclorrhapha. Volumes 11 to X will be divided into parts of convenient size, but it is not possible to specify in advance the taxonomic content of each part. Conciseness and cheapness are main objectives in this new series, and each part will be the work of a specialist, or of a group of specialists. -

Catalogue of Afghanistan Longhorn Beetles (Coleoptera, Cerambycidae) with Two Descriptions of New Phytoecia (Parobereina Danilevsky, 2018) from Central Asia
Humanity space International almanac VOL. 8, No 2, 2019: 104-140 http://zoobank.org/urn:lsid:zoobank.org:pub:30F6FA0A-2D7A-4ED2-9EAE-AB7707FFBE61 Catalogue of Afghanistan Longhorn beetles (Coleoptera, Cerambycidae) with two descriptions of new Phytoecia (Parobereina Danilevsky, 2018) from Central Asia M.A. Lazarev State Budget Professional Educational Institution of the Moscow Region “Chekhov technical college” Novaya str., 4, Novyi Byt village, Chekhov District, Moscow Region 142322 Russia e-mail: [email protected]; [email protected] Key words: Coleoptera, Cerambycidae, taxonomy, distribution, new species, Afghanistan, Pakistan. Abstract: The Catalogue includes all 78 Cerambycidae species of Afghanistan fauna known up to 2019 with the references to the original descriptions; 22 species were not mentioned for Afghanistan in Palaearctic Cerambycidae Catalogue by Löbl & Smetana (2010). Bibliography of each species usually includes the geographical information from corresponding publications. Many new taxonomy positions published after 2010 are used here without special remarks. Agapanthia (Epoptes) dahli ustinovi Danilevsky, 2013 stat. nov. is downgraded from the species level. Two species are described as new Phytoecia (Parobereina) pashtunica sp. n. from Afghanistan and Phytoecia (Parobereina) heinzi sp.n. from Pakistan. The present work is an attempt to summarize all data published up to now on Cerambycidae of Afghanistan fauna. Family CERAMBYCIDAE Latreille, 1802 subfamily Prioninae Latreille, 1802 tribe Macrotomini J. Thomson, 1861 genus Anomophysis Quentin & Villiers, 1981: 374 type species Prionus spinosus Fabricius, 1787 inscripta C.O. Waterhouse, 1884: 380 (Macrotoma) Heyrovský, 1936: 211 - Wama; Tippmann, 1958: 41 - Kabul, Ost- Afghanistan, 1740; Sarobi, am Kabulflus, 900 m; Mangul, Bashgultal, Nuristan, Ost-Afghanistan, 1250 m; Fuchs, 1961: 259 - Sarobi 1100 m, O.-Afghanistan; Fuchs, 1967: 432 - Afghanistan, 25 km N von Barikot, 1800 m, Nuristan; Nimla, 40 km SW von Dschelalabad; Heyrovský, 1967: 156 - Zentral-Afghanistan, Prov. -
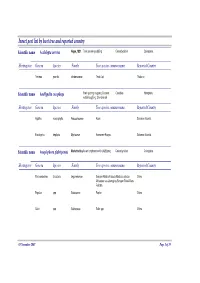
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

Temporal Lags and Overlap in the Diversification of Weevils and Flowering Plants
Temporal lags and overlap in the diversification of weevils and flowering plants Duane D. McKennaa,1, Andrea S. Sequeirab, Adriana E. Marvaldic, and Brian D. Farrella aDepartment of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA 02138; bDepartment of Biological Sciences, Wellesley College, Wellesley, MA 02481; and cInstituto Argentino de Investigaciones de Zonas Aridas, Consejo Nacional de Investigaciones Científicas y Te´cnicas, C.C. 507, 5500 Mendoza, Argentina Edited by May R. Berenbaum, University of Illinois at Urbana-Champaign, Urbana, IL, and approved March 3, 2009 (received for review October 22, 2008) The extraordinary diversity of herbivorous beetles is usually at- tributed to coevolution with angiosperms. However, the degree and nature of contemporaneity in beetle and angiosperm diversi- fication remain unclear. Here we present a large-scale molecular phylogeny for weevils (herbivorous beetles in the superfamily Curculionoidea), one of the most diverse lineages of insects, based on Ϸ8 kilobases of DNA sequence data from a worldwide sample including all families and subfamilies. Estimated divergence times derived from the combined molecular and fossil data indicate diversification into most families occurred on gymnosperms in the Jurassic, beginning Ϸ166 Ma. Subsequent colonization of early crown-group angiosperms occurred during the Early Cretaceous, but this alone evidently did not lead to an immediate and ma- jor diversification event in weevils. Comparative trends in weevil diversification and angiosperm dominance reveal that massive EVOLUTION diversification began in the mid-Cretaceous (ca. 112.0 to 93.5 Ma), when angiosperms first rose to widespread floristic dominance. These and other evidence suggest a deep and complex history of coevolution between weevils and angiosperms, including codiver- sification, resource tracking, and sequential evolution. -

Coleópteros Del Parque Natural De Las Fragas Del Eume
Coleópteros del Parque Natural de las Fragas del Eume (Galicia, noroeste de la Península Ibérica), II: Scarabaeoidea, Buprestoidea, Byrrhoidea, Elateroidea, Bostrichoidea, Lymexyloidea, Cleroidea, Cucujoidea, Tenebrionoidea, Chrysomeloidea y Curculionoidea ANDRÉS BASELGA y FRANCISCO NOVOA Departamento de Biología Animal. Facultad de Biología. Universidad de Santiago de Compostela. 15706 Santiago de Compostela. España. Recibido: 1-VI-2003. Aceptado: 27-I-2004 ISSN: 0210-8984 RESUMEN Se realiza un inventario de 286 especies de Scarabaeoidea, Buprestoidea, Byrrhoidea, Elateroidea, Bostrichoidea, Lymexyloidea, Cleroidea, Cucujoidea, Tenebrionoidea, Chrysomeloidea y Curculionoidea del Parque Natural de las Fragas del Eume (Galicia, noroeste de la Península Ibérica). Destacan especialmente las citas de Aphthona sandrae Baselga y Novoa, 2002 que fue descrita del Parque Natural, así como Chrysobyrrhulus asturiensis Allemand, 1997, Grynobius planus (Fabricius, 1787), Phyllotreta exclamationis (Thunberg, 1784) y Longitarsus fulgens (Foudras, 1860) por ampliar significativamente su distribución. Debido al carácter atlántico del clima y la vegetación, los elementos septentrionales (25,1%) suponen una proporción mucho mayor que los mediterráneos (7,8%). Los endemismos ibéricos son bastan- te abundantes (14,8%) y aparecen numerosas especies típicas de áreas montañosas, datos que indican un alto grado de conservación de esta zona de baja altitud. Palabras clave: Coleoptera, Fragas del Eume, Galicia, Península Ibérica, faunística, biogeografía. ABSTRACT