Inhibition of Mast Cells: a Novel Mechanism by Which Nintedanib
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Prescription Drugs Requiring Prior Authorization
PRESCRIPTION DRUGS REQUIRING PRIOR AUTHORIZATION Revised 10/16 As part of our drug utilization management program, members must request and receive prior authorization for certain prescription drugs in order to use their prescription drug benefits. Below is a list of drugs that currently require prior authorization. This list will be updated periodically as new drugs that require prior authorization are introduced. As benefits may vary by group and individual plans, the inclusion of a medication on this list does not imply prescription drug coverage. The Schedule of Benefits contains a list of drug categories that require prior authorization. Prior authorization requests are processed by our pharmacy benefit manager, Express Scripts, Inc. (ESI). Physicians must call ESI to obtain an authorization. (1-800-842-2015). Drug Name Generic Name Drug Classification Abstral fentanyl citrate oral tablet Controlled Dangerous Substance Accu-Chek Test Strips blood glucose test strips Blood Glucose Test Strips Actemra tocilizumab Monoclonal Antibody Acthar corticotropin Hormone Actimmune interferon gamma 1b Interferon Actiq fentanyl citrate OTFC Controlled Dangerous Substance Adcirca tadalafil Pulmonary Vasodilator Adempas riociguat Pulmonary Vasodilator Adlyxin lixisenatide Type 2 Diabetes Advocate Test Strips blood glucose test strips Blood Glucose Test Strips Aerospan** flunisolide Corticosteroids (Inhaled) Afrezza insulin Insulin (inhaled) Ampyra dalfampridine Multiple Sclerosis Agent Altoprev** lovastatin Cholesterol Alvesco** ciclesonide Corticosteroids -
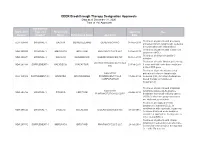
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

CELEBRATING 25 YEARS of PROGRESS for PATIENTS for PROGRESS of YEARS 25 CELEBRATING Annual Report 2019 Report Annual
CELEBRATING 25 YEARS OF PROGRESS FOR PATIENTS FOR PROGRESS YEARS OF 25 CELEBRATING Annual Report 2019 Annual Report Exelixis, Inc. Annual Report 2019 2019 HIGHLIGHTS In 2019, Exelixis marked its 25th anniversary. Our progress during the year highlights how far we’ve come since our early days. More than that, though, it underscores our commitment to pushing scientific and technological boundaries to advance our mission to help cancer patients recover stronger and live longer. As we move through 2020, our team has the same drive and passion for the work as we did when we first opened our doors back in 1994. 610+ employees committed to our mission at year-end 2019, an increase of 27% year over year 12 ongoing or planned trials with label-enabling potential, including four announced in 2019 $1 billion in annual global net revenues for the cabozantinib franchise1 20 ongoingon preclinical programs across ExelixisExxelixis internal disdiscovery and our partners, includingincludi three that ccocouldould reach the clinic this year 4 Commerciallyommerciallymmerciallymerciallyerciallyrciallyciallyiallyallyllyy availablavailablea pprproproduprodproductproducts benefittingttingtingingngg patientsatientstientsientsentsntstlblbi ono a ggloglobal basis 1 Including revenues generated by Ipsen Pharma SAS, our partner in cabozantinib’s global development and commercialization outside of the United States and Japan TO OUR STOCKHOLDERS Before I reflect on our progress in 2019, I want to take a moment to acknowledge the obvious: the coronavirus pandemic is making 2020 a markedly different year for the world, our industry, and for Exelixis. Despite this, I’m confident in our ability to navigate these difficult times and the significant challenges posed by COVID-19. For patients with cancer, continuity of treatment is paramount. -

Your Pharmacy Benefits Welcome to Your Pharmacy Plan
Your Pharmacy Benefits Welcome to Your Pharmacy Plan OptumRx manages your pharmacy benefits for your health plan sponsor. We look forward to helping you make informed decisions about medicines for you and your family. Please review this information to better understand your pharmacy benefit plan. Using Your Member ID Card You can use your pharmacy benefit ID card at any pharmacy participating in your plan’s pharmacy network. Show your ID card each time you fill a prescription at a network pharmacy. They will enter your card information and automatically file it with OptumRx. Your pharmacy will then collect your share of the payment. Your Pharmacy Network Your plan’s pharmacy network includes more than 67,000 independent and chain retail pharmacies nationwide. To find a network pharmacy near you, use the Locate a Pharmacy tool at www.optumrx.com. Or call Customer Service at 1-877-559-2955. Using Non -Network Pharmacies If you do not show the pharmacy your member ID card, or you fill a prescription at a non-network pharmacy, you must pay 100 percent of the pharmacy’s price for the drug. If the drug is covered by your plan, you can be reimbursed. To be reimbursed for eligible prescriptions, simply fill out and send a Direct Member Reimbursement Form with the pharmacy receipt(s) to OptumRx. Reimbursement amounts are based on contracted pharmacy rates minus your copayment or coinsurance. For a copy of the form, go to www.optumrx.com. Or call the Customer Service number above. The number is also on the back of your ID card. -

Australian Public Assessment Report for Nintedanib Esilate
Australian Public Assessment Report for Nintedanib esilate Proprietary Product Name: Ofev and Vargatef Sponsor: Boehringer Ingelheim Pty Ltd February 2016 Therapeutic Goods Administration About the Therapeutic Goods Administration (TGA) • The Therapeutic Goods Administration (TGA) is part of the Australian Government Department of Health and is responsible for regulating medicines and medical devices. • The TGA administers the Therapeutic Goods Act 1989 (the Act), applying a risk management approach designed to ensure therapeutic goods supplied in Australia meet acceptable standards of quality, safety and efficacy (performance) when necessary. • The work of the TGA is based on applying scientific and clinical expertise to decision- making, to ensure that the benefits to consumers outweigh any risks associated with the use of medicines and medical devices. • The TGA relies on the public, healthcare professionals and industry to report problems with medicines or medical devices. TGA investigates reports received by it to determine any necessary regulatory action. • To report a problem with a medicine or medical device, please see the information on the TGA website <https://www.tga.gov.au>. About AusPARs • An Australian Public Assessment Report (AusPAR) provides information about the evaluation of a prescription medicine and the considerations that led the TGA to approve or not approve a prescription medicine submission. • AusPARs are prepared and published by the TGA. • An AusPAR is prepared for submissions that relate to new chemical entities, generic medicines, major variations and extensions of indications. • An AusPAR is a static document; it provides information that relates to a submission at a particular point in time. • A new AusPAR will be developed to reflect changes to indications and/or major variations to a prescription medicine subject to evaluation by the TGA. -

Nintedanib (OFEV) Information for Patients
Form: D-8580 Nintedanib (OFEV) Information for patients Read this medicine fact sheet to learn about: • What this medicine is • Who it is for • How to take it • What side effects to watch for What is nintedanib? Nintedanib [pronounced nin-ten-da-nib] is in a group of medicines called tyrosine-kinase inhibitors and anti-fibrotic agents. Nintedanib affects your body’s immune system and reduces the amount of fibrosis (scarring) in the lungs. Nintedanib is one of two medicines that are approved in Canada to treat Idiopathic Pulmonary Fibrosis (IPF). Another name for this medicine is OFEV. Is nintedanib right for me? Your doctor recommends this medicine for your treatment. But, some patients should not take nintendanib. Tell your doctor: • If you are allergic to nintendanib or any other medicines • If you have an allergy to peanuts • If have an allergy to soy products • If you have ever had serious liver disease or kidney disease. Patients on dialysis should not use nintedanib • If you are pregnant What do I need to know when taking this medicine? Nintedanib can interact with other medicines and health products. This means it might not work well with these products. Tell your doctor: • About all the medicines, supplements and herbal products you are taking. Tell your pharmacist too. • If you are taking erythromycin (Biaxin), ketoconazole, rifampicin (Rifadin, Rofact), phenytoin (Dilantin), St. John’s wart or carbamazepine (Tegretol). You should not take these medicines while you are taking nintedanib. • If you have ever had serious heart disease and high blood pressure. • If you have ever had bleeding or clotting problems. -

Phase II Trial of Nintedanib in Patients with Advanced Esophagogastric Cancer PROTOCOL FACE PAGE for MSKCC THERAPEUTIC/DIAGNOSTIC PROTOCOL
Memorial Sloan Kettering Cancer Center IRB#: 14-094 A (5) Approved: 27-DEC-2016 Phase II trial of Nintedanib in patients with advanced esophagogastric cancer PROTOCOL FACE PAGE FOR MSKCC THERAPEUTIC/DIAGNOSTIC PROTOCOL Principal Yelena Y. Janjigian, MD Medicine Investigator/Department: Co-Principal David H. Ilson, MD, PhD Medicine Investigator(s)/Department: Elizabeth S. Won, MD Medicine Investigator(s)/Department: Marinela Capanu, PhD Epidemiology/Biostatistics Robert Lefkowitz, MD Radiology Marc Z. Simmons, MD Radiology Ghassan Abou-Alfa, MD Medicine Andrea Cercek, MD Medicine Andrew Epstein, MD Medicine James Harding, MD Medicine David Kelsen, MD Medicine Nancy Kemeny, MD Medicine Geoffrey Ku, MD Medicine Maeve Lowery, MD Medicine Eileen O’Reilly, MD Medicine Diane Reidy-Lagunes, MD Medicine Leonard Saltz, MD Medicine Neil Segal, MD, PhD Medicine Zsofia Stadler, MD Medicine Kenneth H. Yu, MD Medicine Anna Varghese, MD Medicine Rona Yaeger, MD Medicine Nitya Raj, MD Medicine Ellen Hollywood, NP Nursing Maria Pacis, NP Nursing Erica Kaufmann, NP Nursing Sree Chalasani, MD Medicine Han Xiao, MD Medicine Afsheen Iqbal, MD Medicine Leticia Smith, APN Nursing Janet Cogswell, CRN Nursing Avni Desai, MD Medicine John Fiore, MD Medicine Stuart Lichtman, MD Medicine Jahan Aghalar, MD Medicine Juliana Eng, MD Medicine Jia Li, MD Medicine Wanqing Iris Zhi, MD, PhD Medicine Adriana Olivo, NP Nursing Lori Gofter, CRN Nursing Arlyn Apollo, MD Medicine Page 1 of 36 Memorial Sloan Kettering Cancer Center IRB#: 14-094 A (5) Approved: 27-DEC-2016 Pamela Drullinsky, MD Medicine Zoe Goldberg, MD Medicine Tiffany Troso-Sandoval, MD Medicine Kenneth Ng, MD Medicine Erin Scansarole, CRN Nursing Krysti Corrado, CRN Nursing Philip Caron, MD Medicine Alice Zervoudakis, MD Medicine Parisa Momtaz, MD Medicine Chung-Han Lee, MD Medicine Gloria Wasilewski, CRN Nursing Jason Konner, MD Medicine Serena Wong, MD Medicine Jacqueline Bromberg, MD Medicine PhD Colette Owens, MD Medicine Loren Michel, MD Medicine Azadeh Namakydoust, MD Medicine Marina Shcherba, DO Medicine Consenting Yelena Y. -

Use of Multitarget Tyrosine Kinase Inhibitors to Attenuate Platelet-Derived Growth Factor Signalling in Lung Disease
REVIEW PDGF SIGNALLING Use of multitarget tyrosine kinase inhibitors to attenuate platelet-derived growth factor signalling in lung disease Rana Kanaan and Charlie Strange Affiliation: Division of Pulmonary, Critical Care, Allergy and Sleep Medicine, Dept of Medicine, Medical University of South Carolina, Charleston, SC, USA. Correspondence: Charlie Strange, MSC630, Medical University of South Carolina, Charleston, SC 29425, USA. E-mail: [email protected] @ERSpublications An increasing role for PDGF signalling inhibitors in clinical trials for the treatment of various pulmonary diseases http://ow.ly/buaI30f9HcN Cite this article as: Kanaan R, Strange C. Use of multitarget tyrosine kinase inhibitors to attenuate platelet- derived growth factor signalling in lung disease. Eur Respir Rev 2017; 26: 170061 [https://doi.org/10.1183/ 16000617.0061-2017]. ABSTRACT Platelet-derived growth factors (PDGFs) and their receptors (PDGFRs) play a fundamental role in the embryonic development of the lung. Aberrant PDGF signalling has been documented convincingly in a large variety of pulmonary diseases, including idiopathic pulmonary arterial hypertension, lung cancer and lung fibrosis. Targeting PDGF signalling has been proven to be effective in these diseases. In clinical practice, the most effective way to block PDGF signalling is to inhibit the activity of the intracellular PDGFR kinases. Although the mechanism of action of such drugs is not specific for PDGF signalling, the medications have a broad therapeutic index that allows clinical use. The safety profile and therapeutic opportunities of these and future medications that target PDGFs and PDGFRs are reviewed. Introduction Platelet-derived growth factors (PDGFs) and their receptors (PDGFRs) represent one of the most intensively studied families of signalling factors over the past four decades. -

Nintedanib Ameliorates Experimental Pulmonary Arterial
bioRxiv preprint doi: https://doi.org/10.1101/584110; this version posted March 20, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. 1 Nintedanib ameliorates experimental pulmonary arterial hypertension via 2 inhibition of endothelial mesenchymal transition and smooth muscle cell proliferation 3 4 5 Takeo Tsutsumi1, Tetsutaro Nagaoka1, Takashi Yoshida1, Lei Wang1, 6 Sachiko Kuriyama1, Yoshifumi Suzuki1, Yuichi Nagata1, 7 Norihiro Harada1, Yuzo Kodama1, Fumiyuki Takahashi1, Yoshiteru Morio2, 8 and Kazuhisa Takahashi1. 9 10 1 Department of Respiratory Medicine, Juntendo University Faculty of Medicine & Graduate 11 School of Medicine, 3-1-3 Hongo Bunkyo-ku, Tokyo 113-8431, Japan. 12 2 Department of Respiratory Medicine, National Hospital Organization Tokyo National 13 Hospital, 3-1-1 Takeoka Kiyose-shi, Tokyo 204-8585, Japan. 14 15 Short title: 16 Nintedanib for experimental pulmonary arterial hypertension 17 18 Corresponding Author: 19 Tetsutaro Nagaoka MD PhD 20 Department of Respiratory Medicine, Juntendo University Graduate School of Medicine 21 3-1-3 Hongo Bunkyo-ku, Tokyo 113-8431, Japan. 22 Tel: +81-3-5802-1063 23 Fax: +81-3-5802-1617 24 E-mail: [email protected] 25 26 Author contributions: 27 T.T contributed whole of experiment. T.N supervised whole of this study. T.Y, L.W and S.K 28 assisted in molecular biological experiment. Y.S and Y.N assisted in animal experiment. 29 N.H, Y.K, F.T, Y.M, and K.T assisted in planning of this study. -

HIGHLIGHTS of PRESCRIBING INFORMATION Thereafter Or As Clinically Indicated
HIGHLIGHTS OF PRESCRIBING INFORMATION thereafter or as clinically indicated. Temporary dosage reductions or These highlights do not include all the information needed to use OFEV discontinuations may be required. (2.1, 2.3, 5.2) safely and effectively. See full prescribing information for OFEV. Gastrointestinal disorders: Diarrhea, nausea, and vomiting have occurred with OFEV. Treat patients at first signs with adequate hydration and OFEV® (nintedanib capsules), for oral use antidiarrheal medicine (e.g., loperamide) or anti-emetics. Discontinue Initial U.S. Approval: 2014 OFEV if severe diarrhea, nausea, or vomiting persists despite symptomatic treatment. (5.3) ---------------------------RECENT MAJOR CHANGES--------------------------- Embryo-Fetal toxicity: Can cause fetal harm. Advise females of Indications and Usage (1) 3/2020 reproductive potential of the potential risk to a fetus and to use highly Warnings and Precautions (5) 10/2020 effective contraception. Advise women taking oral hormonal contraceptives experiencing vomiting, diarrhea, or other conditions where ----------------------------INDICATIONS AND USAGE--------------------------- the drug absorption may be reduced to use alternative highly effective OFEV is a kinase inhibitor indicated for: contraception. (5.4, 8.1, 8.3) Treatment of idiopathic pulmonary fibrosis (IPF). (1.1) Arterial thromboembolic events have been reported. Use caution when Treatment of chronic fibrosing interstitial lung diseases (ILDs) with a treating patients at higher cardiovascular risk including known coronary progressive phenotype (1.2) artery disease. (5.5) Slowing the rate of decline in pulmonary function in patients with systemic Bleeding events have been reported. Use OFEV in patients with known sclerosis-associated interstitial lung disease (SSc-ILD). (1.3) bleeding risk only if anticipated benefit outweighs the potential risk. -

Lorlatinib (PF-06463922) for Advanced ALK Or ROS1 Positive Non-Small Cell Lung Cancer – Second Line NIHRIO (HSRIC) ID: 14896 NICE ID: 9656
NIHR Innovation Observatory Evidence Briefing: August 2017 Lorlatinib (PF-06463922) for advanced ALK or ROS1 positive non-small cell lung cancer – second line NIHRIO (HSRIC) ID: 14896 NICE ID: 9656 LAY SUMMARY Lung cancer is the second most common type of cancer. The most common type of lung cancer is non- small cell lung cancer (NSCLC). NSCLC has three sub-types that can be further grouped by their specific genetic mutation. Two of these types are ALK-positive and ROS1-positive lung cancer, both of which fuse with other genes and promote cancer tumour growth. Patients with ALK-positive and ROS1-positive NSCLC are similar in that they are often younger, have no history of smoking and have a particular type of NSCLC called adenocarcinoma (a cancerous tumour of the lung). Lorlatinib is a new drug under development for the sub-group of advanced non-small cell lung cancer who are ALK or ROS1 positive and have already undergone gene treatment with drugs that specifically target this type of cancer. Lorlatinib is currently being evaluated in phase II clinical trials as an oral dose at 100 mg daily. If marketed it will become an additional targeted treatment for this sub-group of ALK or ROS1 positive patients. This briefing is based on information available at the time of research and a limited literature search. It is not intended to be a definitive statement on the safety, efficacy or effectiveness of the health technology covered and should not be used for commercial purposes or commissioning without additional information. This briefing presents independent research funded by the National Institute for Health Research (NIHR). -

The Emerging Therapeutic Landscape of ALK Inhibitors in Non-Small Cell Lung Cancer
pharmaceuticals Review The Emerging Therapeutic Landscape of ALK Inhibitors in Non-Small Cell Lung Cancer 1, 1, 1, 1 Valerio Gristina y, Maria La Mantia y , Federica Iacono y, Antonio Galvano , 1, 2, , Antonio Russo z and Viviana Bazan * z 1 Department of Surgical, Oncological and Oral Sciences, University of Palermo, 90127 Palermo, Italy; [email protected] (V.G.); [email protected] (M.L.M.); [email protected] (F.I.); [email protected] (A.G.); [email protected] (A.R.) 2 Department of Biomedicine, Neuroscience and Advanced Diagnostics (Bi.N.D.), Section of Medical Oncology, University of Palermo, 90127 Palermo, Italy * Correspondence: [email protected] These authors contributed equally to this work. y These authors are co-last. z Received: 29 October 2020; Accepted: 16 December 2020; Published: 18 December 2020 Abstract: The treatment of metastatic non-small cell lung cancer (NSCLC) has undergone a paradigm shift over the last decade. Better molecular characterization of the disease has led to the rapid improvement of personalized medicine and the prompt delivery of targeted therapies to patients with NSCLC. The discovery of the EML4-ALK fusion gene in a limited subset of patients affected by NSCLC and the subsequent clinical development of crizotinib in 2011 has been an impressive milestone in lung cancer research. Unfortunately, acquired resistances regularly develop, hence disease progression occurs. Afterward, modern tyrosine kinase inhibitors (TKIs), such as ceritinib, alectinib, brigatinib, and lorlatinib, have been approved by the Food and Drug Administration (FDA) for the management of anaplastic lymphoma kinase (ALK)-positive NSCLCs.