Living Fossils Among Tiny Allogastropoda with High and Slender Shell from the Reef Environment of the Gulf of Aqaba with Remarks on Fossil and Recent Relatives
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Caenogastropoda Eulimidae) from the Western Iberian Peninsula
Biodiversity Journal, 2021, 12 (2): 277–282, https://doi.org/10.31396/Biodiv.Jour.2021.12.2.277.282 https://zoobank.org:pub:AA55BDF3-1E5E-469D-84A8-5EC6A013150F A new minute eulimid (Caenogastropoda Eulimidae) from the western Iberian Peninsula Serge Gofas1 & Luigi Romani2* 1Departamento de Biología Animal, Universidad de Málaga, Campus de Teatinos s/n, 29071 Málaga, Spain,; e-mail: [email protected] 2Via delle ville 79, 55012 Capannori (Lucca), Italy; e-mail: [email protected] *Corresponding author ABSTRACT An enigmatic small-sized gastropod is recorded on few shells originating from the western Iberian Peninsula. It is assigned to the family Eulimidae relying on shell characters, and com- pared to species of several genera which share some morphological features with it. It is de- scribed as new and provisionally included in Chileutomia Tate et Cossmann, 1898, although with reservation, as we refrain to establish a new genus without anatomical and molecular data which can clarify the phylogenetic relationships of the new species. KEY WORDS Gastropoda; new species; NW Atlantic Ocean. Received 06.01.2020; accepted 28.02.2021; published online 12.04.2021 INTRODUCTION tematics and intra-familial relationships is at its very beginning, for instance the phylogenetic posi- The Eulimidae Philippi, 1853 are a species-rich tion of the Eulimidae within the Caenogastropoda taxon of marine snails, mostly parasitic of Echino- was assessed by molecular means only recently dermata (Warén, 1984). The family comprises (Takano & Kano, 2014), leading to consider them about one thousand recent valid species recognized as sister-group to the Vanikoridae (Bouchet et al., worldwide (MolluscaBase, 2021a), but a more re- 2017). -

Deep-Sea Fauna of the European Seas: an Annotated Species Check-List Of
Invertebrate Zoology, 2014, 11(1): 134–155 © INVERTEBRATE ZOOLOGY, 2014 Deep-sea fauna of European seas: An annotated species check-list of benthic invertebrates living deeper than 2000 m in the seas bordering Europe. Gastropoda Alexander V. Sysoev Zoological Museum, Moscow State University, Bol’shaya Nikitskaya ul., 6, Moscow, 125009, Russia. E-mail: [email protected] ABSTRACT: An annotated check-list is given of Gastropoda species occurring deeper than 2000 m in the seas bordering Europe. The check-list is based on published data. The check- list includes 221 species. For each species data on localities in European seas and general species distribution are provided. Station data are presented separately in the present thematic issue. How to cite this article: Sysoev A.V. 2014. Deep-sea fauna of European seas: An annotated species check-list of benthic invertebrates living deeper than 2000 m in the seas bordering Europe. Gastropoda // Invert. Zool. Vol.11. No.1. P.134–155. KEY WORDS: deep-sea fauna, European seas, Gastropoda. Глубоководная фауна европейских морей: аннотированный список видов донных беспозвоночных, обитающих глубже 2000 м в морях, окружающих Европу. Gastropoda А.В. Сысоев Зоологический музей МГУ им. М.В. Ломоносова, ул. Большая Никитская, 6, Москва 125009, Россия. E-mail: [email protected] РЕЗЮМЕ: Приводится аннотированный список видов Gastropoda, обитающих глуб- же 2000 м в морях, окружающих Европу. Список основан на опубликованных данных. Список насчитывает 221 вид. Для каждого вида приведены данные о нахождениях в европейских морях и сведения о распространении. Данные о станци- ях приводятся в отдельном разделе настоящего тематического выпуска. Как цитировать эту статью: Sysoev A.V. -

Descripción De Nuevas Especies Animales De La Península Ibérica E Islas Baleares (1978-1994): Tendencias Taxonómicas Y Listado Sistemático
Graellsia, 53: 111-175 (1997) DESCRIPCIÓN DE NUEVAS ESPECIES ANIMALES DE LA PENÍNSULA IBÉRICA E ISLAS BALEARES (1978-1994): TENDENCIAS TAXONÓMICAS Y LISTADO SISTEMÁTICO M. Esteban (*) y B. Sanchiz (*) RESUMEN Durante el periodo 1978-1994 se han descrito cerca de 2.000 especies animales nue- vas para la ciencia en territorio ibérico-balear. Se presenta como apéndice un listado completo de las especies (1978-1993), ordenadas taxonómicamente, así como de sus referencias bibliográficas. Como tendencias generales en este proceso de inventario de la biodiversidad se aprecia un incremento moderado y sostenido en el número de taxones descritos, junto a una cada vez mayor contribución de los autores españoles. Es cada vez mayor el número de especies publicadas en revistas que aparecen en el Science Citation Index, así como el uso del idioma inglés. La mayoría de los phyla, clases u órdenes mues- tran gran variación en la cantidad de especies descritas cada año, dado el pequeño núme- ro absoluto de publicaciones. Los insectos son claramente el colectivo más estudiado, pero se aprecia una disminución en su importancia relativa, asociada al incremento de estudios en grupos poco conocidos como los nematodos. Palabras clave: Biodiversidad; Taxonomía; Península Ibérica; España; Portugal; Baleares. ABSTRACT Description of new animal species from the Iberian Peninsula and Balearic Islands (1978-1994): Taxonomic trends and systematic list During the period 1978-1994 about 2.000 new animal species have been described in the Iberian Peninsula and the Balearic Islands. A complete list of these new species for 1978-1993, taxonomically arranged, and their bibliographic references is given in an appendix. -

The Ultrastructure of Spermatozoa and Spermiogenesis in Pyramidellid Gastropods, and Its Systematic Importance John M
HELGOLANDER MEERESUNTERSUCHUNGEN Helgol~inder Meeresunters. 42,303-318 (1988) The ultrastructure of spermatozoa and spermiogenesis in pyramidellid gastropods, and its systematic importance John M. Healy School of Biological Sciences (Zoology, A08), University of Sydney; 2006, New South Wales, Australia ABSTRACT: Ultrastructural observations on spermiogenesis and spermatozoa of selected pyramidellid gastropods (species of Turbonilla, ~gulina, Cingufina and Hinemoa) are presented. During spermatid development, the condensing nucleus becomes initially anterio-posteriorly com- pressed or sometimes cup-shaped. Concurrently, the acrosomal complex attaches to an electron- dense layer at the presumptive anterior pole of the nucleus, while at the opposite (posterior) pole of the nucleus a shallow invagination is formed to accommodate the centriolar derivative. Midpiece formation begins soon after these events have taken place, and involves the following processes: (1) the wrapping of individual mitochondria around the axoneme/coarse fibre complex; (2) later internal metamorphosis resulting in replacement of cristae by paracrystalline layers which envelope the matrix material; and (3) formation of a glycogen-filled helix within the mitochondrial derivative (via a secondary wrapping of mitochondria). Advanced stages of nuclear condensation {elongation, transformation of fibres into lamellae, subsequent compaction) and midpiece formation proceed within a microtubular sheath ('manchette'). Pyramidellid spermatozoa consist of an acrosomal complex (round -

Turritella Communis (Risso, 1826)
ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΓΕΩΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΙΚΗΣ ΘΑΛΑΣΣΙΑΣ ΓΕΩΛΟΓΙΑΣ ΚΑΙ ΓΕΩΔΥΝΑΜΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΘΑΛΑΣΣΙΑΣ ΓΕΩΛΟΓΙΑΣ ΚΑΙ ΦΥΣΙΚΗΣ ΩΚΕΑΝΟΓΡΑΦΙΑΣ ΜΙΚΡΟΠΑΛΑΙΟΝΤΟΛΟΓΙΚΕΣ ΑΝΑΛΥΣΕΙΣ ΘΑΛΑΣΣΙΩΝ ΙΖΗΜΑΤΩΝ ΤΟΥ ΑΜΒΡΑΚΙΚΟΥ ΚΟΛΠΟΥ Μεταπτυχιακή Φοιτήτρια Αρκαδιανού Μαρία Επιβλέπουσα Καθηγήτρια: Γεραγά Μαρία Τριμελής Επιτροπή: Γεραγά Μαρία Παπαθεοδώρου Γεώργιος Ηλιόπουλος Γεώργιος 0 Πάτρα, 2019 Αφιερώνεται στην οικογένειά μου που με στηρίζει σε κάθε νέο ξεκίνημα σε κάθε απόφαση… Ευχαριστίες… Θα ήθελα να ευχαριστήσω τους καθηγητές Γεραγά Μαρία και Παπαθεοδώρου Γεώργιο που μου έδωσαν τη δυνατότητα να εκπονήσω τη διπλωματική μου εργασία στον τομέα της Περιβαλλοντικής Ωκεανογραφίας δείχνοντάς μου εμπιστοσύνη και προσφέροντάς μου πολύτιμη βοήθεια. Τον καθηγητή μου Ηλιόπουλο Γεώργιο και την Δρ. Παπαδοπούλου Πηνελόπη για την πολύτιμη βοήθεια τους, την καθοδήγηση, τις χρήσιμες συμβουλές και τη στήριξη. Την Υποψήφια Διδάκτορα Τσώνη Μαρία για τις συμβουλές της στο χειρισμό ορισμένων λογισμικών. Την Υποψήφια Διδάκτορα Πρανδέκου Αμαλία για τις συμβουλές της. Τον φίλο μου Ανδρέα για τη στήριξη και την υπομονή του. Τέλος, θέλω να εκφράσω ένα μεγάλο ευχαριστώ στην οικογένειά μου που με στήριξε με κάθε τρόπο σε όλη τη διάρκεια των σπουδών μου και υπήρξαν σημαντικοί αρωγοί όλα αυτά τα χρόνια. 1 Ευχαριστίες……………………………………………………………………………………………………….....................1 ΕΙΣΑΓΩΓΗ………………………………………………………………………………………………………………………………3 Σκοπός…………………………………………………………………………………………………..................3 1. ΠΕΡΙΟΧΗ ΜΕΛΕΤΗΣ………………………………………………………………………………………………3 1.1Γενικά …………………………………………………………………………………………………………………3 -

Faunal Change and Bathymetric Diversity Gradient in Deep-Sea Prosobranchs from Northeastern Atlantic
Biodiversity and Conservation (2006) Ó Springer 2006 DOI 10.1007/s10531-005-1344-9 -1 Faunal change and bathymetric diversity gradient in deep-sea prosobranchs from Northeastern Atlantic CELIA OLABARRIA1,2 1Southampton Oceanography Centre, DEEPSEAS Benthic Biology Group, Empress Dock, South- ampton SO14 3ZH, United Kingdom; 2Present address: Departamento de Ecoloxı´a e Bioloxı´a Animal, Area Ecoloxı´a, Universidad de Vigo, Campus Lagoas-Marcosende, 36310 Vigo (Pontevedra), Spain (e-mail: [email protected]; phone: +34-986-812587; fax: +34-986-812556) Received 6 January 2005; accepted in revised form 11 July 2005 Key words: Deep sea, Diversity, Faunal turnover, Northeastern Atlantic, Porcupine Abyssal Plain, Porcupine Seabight, Prosobranchs Abstract. Despite the plethora of studies, geographic patterns of diversity in deep sea remain subject of speculation. This study considers a large dataset to examine the faunal change and depth-diversity gradient of prosobranch molluscs in the Porcupine Seabight and adjacent Abyssal Plain (NE Atlantic). Rates of species succession (addition and loss) increased rapidly with increasing depth and indicated four possible areas of faunal turnover at about 700, 1600, 2800 and 4100 m. Depth was a significant predictor of diversity, explaining nearly a quarter the variance. There was a pattern of decreasing diversity downslope from 250 m to 1500–1600 m, followed by an increase to high values at about 4000 m and then again, a fall to 4915 m. Processes causing diversity patterns of prosobranchs in the Porcupine Seabight and adjacent Abyssal Plain are likely to differ in magnitude or type, from those operating in other Atlantic areas. Introduction An increasing focus for biodiversity research in the deep sea has been to test for the existence of large-scale gradients in the diversity of marine soft- sediment fauna in deep sea (e.g. -

THE FESTIVUS Page Iii
ISSN 0738-9388 ISSN 0738-9388 TH E F E S T I VU S T H E F E S T I VU S A publication of the San Diego Shell Club A publication of the San Diego Shell Club Volume XXXIX September 9, 2007 Supplement Volume XXXIX September 9, 2007 Supplement The Recent Molluscan Fauna of Île Clipperton The Recent Molluscan Fauna of Île Clipperton (Tropical Eastern Pacific) (Tropical Eastern Pacific) Kirstie L. Kaiser Kirstie L. Kaiser US US Vol. XXXIX: Supplement THE FESTIV Page i Vol. XXXIX: Supplement THE FESTIV Page i ISSN 0738-9388 ISSN 0738-9388 THE RECENT MOLLUSCAN FAUNA OF ÎLE CLIPPERTON THE RECENT MOLLUSCAN FAUNA OF ÎLE CLIPPERTON (TROPICAL EASTERN PACIFIC) (TROPICAL EASTERN PACIFIC) KIRSTIE L. KAISER KIRSTIE L. KAISER Research Associate, Santa Barbara Museum of Natural History Research Associate, Santa Barbara Museum of Natural History 2559 Puesta del Sol Road, Santa Barbara, California 93105, USA 2559 Puesta del Sol Road, Santa Barbara, California 93105, USA Email: [email protected] Email: [email protected] September 9, 2007 September 9, 2007 Front Cover: Sunrise at Clipperton. Looking East-SE across the lagoon to Clipperton Rock. Photograph taken by Camille Front Cover: Sunrise at Clipperton. Looking East-SE across the lagoon to Clipperton Rock. Photograph taken by Camille Fresser on 14 January 2005 at 7:49 a.m. Fresser on 14 January 2005 at 7:49 a.m. Front (inside) Cover: Bathymetric chart of Île Clipperton. Copyright: Septième Continent – Jean-Louis Etienne, Expédition Front (inside) Cover: Bathymetric chart of Île Clipperton. Copyright: Septième Continent – Jean-Louis Etienne, Expédition Clipperton. -

NEAT Mollusca
NEAT (North East Atlantic Taxa): Scandinavian marine Mollusca Check-List compiled at TMBL (Tjärnö Marine Biological Laboratory) by: Hans G. Hansson 1994-02-02 / small revisions until February 1997, when it was published on Internet as a pdf file and then republished August 1998.. Citation suggested: Hansson, H.G. (Comp.), NEAT (North East Atlantic Taxa): Scandinavian marine Mollusca Check-List. Internet Ed., Aug. 1998. [http://www.tmbl.gu.se]. Denotations: (™) = Genotype @ = Associated to * = General note PHYLUM, CLASSIS, SUBCLASSIS, SUPERORDO, ORDO, SUBORDO, INFRAORDO, Superfamilia, Familia, Subfamilia, Genus & species N.B.: This is one of several preliminary check-lists, covering S Scandinavian marine animal (and partly marine protoctistan) taxa. Some financial support from (or via) NKMB (Nordiskt Kollegium för Marin Biologi), during the last years of the existence of this organization (until 1993), is thankfully acknowledged. The primary purpose of these checklists is to faciliate for everyone, trying to identify organisms from the area, to know which species that earlier have been encountered there, or in neighbouring areas. A secondary purpose is to faciliate for non-experts to put as correct names as possible on organisms, including names of authors and years of description. So far these checklists are very preliminary. Due to restricted access to literature there are (some known, and probably many unknown) omissions in the lists. Certainly also several errors may be found, especially regarding taxa like Plathelminthes and Nematoda, where the experience of the compiler is very rudimentary, or. e.g. Porifera, where, at least in certain families, taxonomic confusion seems to prevail. This is very much a small modernization of T. -

Influence of Ecological Role on Bathymetric Patterns of Deep-Sea Species: Size Clines in Parasitic Gastropods
MARINE ECOLOGY PROGRESS SERIES Vol. 320: 161–167, 2006 Published August 29 Mar Ecol Prog Ser Influence of ecological role on bathymetric patterns of deep-sea species: size clines in parasitic gastropods Craig R. McClain1, 3,*, Jennifer Crouse2 1Department of Biology, University of New Mexico, Albuquerque, New Mexico 87131, USA 2Department of Biology, University of Massachusetts, 100 Morrissey Blvd., Boston, Massachusetts 02125, USA 3Present address: Monterey Bay Aquarium Research Institute (MBARI), 7700 Sandholdt Road, Moss Landing, California 95039, USA ABSTRACT: How an organism’s phenotype responds to both its biotic and abiotic environment is a complex interplay of selection pressures and adaptive tradeoffs. Bathymetric patterns of body size in deep-sea organisms should also reflect both ecological role and taxon-specific constraints, as exem- plified in a variety of recent studies. Here, we examine bathymetric size clines in deep-sea ectopara- sites, a group that has received little attention in the literature compared to other deep-sea groups. Specifically, we focus on body size in 3 families (21 species) of ptenoglossate gastropods from the deep western North Atlantic, conducting analyses both within and among species. Both quantile and linear regression models yielded non-significant relationships for body size and depth for the 3 most abundant species. Two of the 3 families exhibited positive size–depth relationships, but only in mean size. The findings indicate that resource availability/host size may control parasite density, but dis- lodgement and predation risk may set a hard upper boundary on body size in deep-sea ectoparasites. Moreover, this study stresses the necessity of understanding the ecological role of species in investi- gating body size trends. -
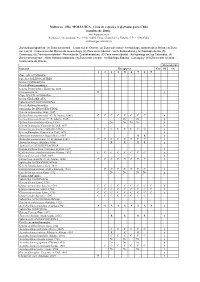
Moluscos - Filo MOLLUSCA
Moluscos - Filo MOLLUSCA. Lista de especies registradas para Cuba (octubre de 2006). José Espinosa Sáez Instituto de Oceanología, Ave 1ª No. 18406, Playa, Ciudad de La Habana, C.P. 11200, Cuba [email protected] Zonas biogeográficas: (1) Zona suroriental – Costa sur de Oriente, (2) Zona surcentral - Archipiélago Jardines de la Reina, (3) Zona sur central - Costa al sur del Macizo de Guamuhaya, (4) Zona suroccidental - Golfo de Batabanó y Archipiélago de los, (5) Canarreos, (6) Zona suroccidental - Península de Guanahacabibes, (7) Zona noroccidental - Archipiélago de Los Colorados, (8) Zona noroccidental - Norte Habana-Matanzas, (9) Zona norte-central - Archipiélago Sabana - Camagüey, (10) Zona norte-oriental - Costa norte de Oriente Abreviaturas Especies Bioegiones Cu Pl Oc 1 2 3 4 5 6 7 8 9 Clase APLACOPHORA Subclase SOLENOGASTRES Orden CAVIBELONIA Familia Proneomeniidae Género Proneomenia Hubrecht, 1880 Proneomenia sp . R x Clase POLYPLACOPHORA Orden NEOLORICATA Suborden ISCHNOCHITONINA Familia Ischnochitonidae Subfamilia ISCHNOCHITONINAE Género Ischnochiton Gray, 1847 Ischnochiton erythronotus (C. B. Adams, 1845) C C C C C C C C x Ischnochiton papillosus (C. B. Adams, 1845) Nc Nc x Ischnochiton striolatus (Gray, 1828) Nc Nc Nc Nc x Género Ischnoplax Carpenter in Dall, 1879 x Ischnoplax pectinatus (Sowerby, 1832) C C C C C C C C x Género Stenoplax Carpenter in Dall, 1879 x Stenoplax bahamensis Kaas y Belle, 1987 R R x Stenoplax purpurascens (C. B. Adams, 1845) C C C C C C C C x Stenoplax boogii (Haddon, 1886) R R R R x Subfamilia CALLISTOPLACINAE Género Callistochiton Carpenter in Dall, 1879 x Callistochiton shuttleworthianus Pilsbry, 1893 C C C C C C C C x Género Ceratozona Dall, 1882 x Ceratozona squalida (C. -

Heterobranchia (Gastropoda) from the Jurassic Deposits of Russia A
ISSN 0031-0301, Paleontological Journal, 2019, Vol. 53, No. 11, pp. 1162–1187. © Pleiades Publishing, Ltd., 2019. Heterobranchia (Gastropoda) from the Jurassic Deposits of Russia A. V. Guzhov* Borissiak Paleontological Institute, Russian Academy of Sciences, Moscow, 117647 Russia *e-mail: [email protected] Received March 11, 2018; revised November 30, 2018; accepted December 28, 2018 Abstract—Small gastropods from the Jurassic deposits of the European Russia, united into the lower Hetero- branchia or Allogastropoda, are described. The families of Ampezzanildidae, Cimidae, Cornirostridae, Ebal- idae, and Stuoraxidae are distinguished, and a set of taxa is given without reference to a definite family. The family Ampezzanildidae from the Jurassic deposits is described for the first time on the basis of the mass material, ascribed to the new genus Zizipupa gen. nov. with the sole species of Z. costata sp. nov. The family Cim- idae includes the genera of Cristalloella, Rotfanella, Urlocella, and Unzhispira gen. nov. with species C. spiralo- costata (Gründel, 1998), R. gerasimovi sp. nov., R. reticulata sp. nov., Urlocella undulata sp. nov., and Unzhispira minuta sp. nov. The genus Heteronatica gen. nov. is included into the family Cornirostridae. This genus is the first siphonostomatous representative of the given family including the sole long-living species H. globosa sp. nov., which is subdivided into subspecies H. globosa globosa and H. globosa promota subsp. nov. The family Ebalidae is represented by the genus Ebala, shells of which are distributed from the Middle Oxfordian to the Middle Volgian. The family Stuoraxidae is described based on two genera Stuoraxis and Aneudaronia gen. nov., including species of S. -

Evolution, Distribution, and Phylogenetic Clumping of a Repeated Gastropod Innovation
Zoological Journal of the Linnean Society, 2017, 180, 732–754. With 5 figures. The varix: evolution, distribution, and phylogenetic clumping of a repeated gastropod innovation NICOLE B. WEBSTER1* and GEERAT J. VERMEIJ2 1Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canada T6G 2E9 2Department of Earth and Planetary Sciences, University of California, Davis, CA 95616, USA Received 27 June 2016; revised 4 October 2016; accepted for publication 25 October 2016 A recurrent theme in evolution is the repeated, independent origin of broadly adaptive, architecturally and function- ally similar traits and structures. One such is the varix, a shell-sculpture innovation in gastropods. This periodic shell thickening functions mainly to defend the animal against shell crushing and peeling predators. Varices can be highly elaborate, forming broad wings or spines, and are often aligned in synchronous patterns. Here we define the different types of varices, explore their function and morphological variation, document the recent and fossil distri- bution of varicate taxa, and discuss emergent patterns of evolution. We conservatively found 41 separate origins of varices, which were concentrated in the more derived gastropod clades and generally arose since the mid-Mesozoic. Varices are more prevalent among marine, warm, and shallow waters, where predation is intense, on high-spired shells and in clades with collabral ribs. Diversification rates were correlated in a few cases with the presence of varices, especially in the Muricidae and Tonnoidea, but more than half of the origins are represented by three or fewer genera. Varices arose many times in many forms, but generally in a phylogenetically clumped manner (more frequently in particular higher taxa), a pattern common to many adaptations.