Implications of a 'Third Signal' in NK Cells
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Primary Sjogren Syndrome: Focus on Innate Immune Cells and Inflammation
Review Primary Sjogren Syndrome: Focus on Innate Immune Cells and Inflammation Chiara Rizzo 1, Giulia Grasso 1, Giulia Maria Destro Castaniti 1, Francesco Ciccia 2 and Giuliana Guggino 1,* 1 Department of Health Promotion, Mother and Child Care, Internal Medicine and Medical Specialties, Rheumatology Section, University of Palermo, Piazza delle Cliniche 2, 90110 Palermo, Italy; [email protected] (C.R.); [email protected] (G.G.); [email protected] (G.M.D.C.) 2 Department of Precision Medicine, University of Campania “Luigi Vanvitelli”, Via L. De Crecchio 7, 80138 Naples, Italy; [email protected] * Correspondence: [email protected]; Tel.: +39-091-6552260 Received: 30 April 2020; Accepted: 29 May 2020; Published: 3 June 2020 Abstract: Primary Sjogren Syndrome (pSS) is a complex, multifactorial rheumatic disease that mainly targets salivary and lacrimal glands, inducing epithelitis. The cause behind the autoimmunity outbreak in pSS is still elusive; however, it seems related to an aberrant reaction to exogenous triggers such as viruses, combined with individual genetic pre-disposition. For a long time, autoantibodies were considered as the hallmarks of this disease; however, more recently the complex interplay between innate and adaptive immunity as well as the consequent inflammatory process have emerged as the main mechanisms of pSS pathogenesis. The present review will focus on innate cells and on the principal mechanisms of inflammation connected. In the first part, an overview of innate cells involved in pSS pathogenesis is provided, stressing in particular the role of Innate Lymphoid Cells (ILCs). Subsequently we have highlighted the main inflammatory pathways, including intra- and extra-cellular players. -

Recombinant Human NCR3/Nkp300 Protein
Leader in Biomolecular Solutions for Life Science Recombinant Human NCR3/NKp300 Protein Catalog No.: RP00179 Recombinant Sequence Information Background Species Gene ID Swiss Prot Natural Cytotoxicity Triggering Receptor 3, NCR3, also known as NKp30, or CD337, Human 259197 O14931 is a natural cytotoxicity receptor. NKp30 is expressed on both resting and activated NK cells of the CD56dim, CD16+ subset that account for more that 85% of NK cells Tags found in peripheral blood and spleen. NKp30 is absent from the CD56bright, CD16- C-Fc & 6×His subset that constitutes the majority of NK cells in lymph node and tonsil, however, its expression is up-regulated in these cells upon IL-2 activation .NKp30 is a Synonyms member of the immunoglobulin superfamily and one of three existing natural 1C7;CD337;DAAP-90L16.3;LY117;MALS cytotoxicity-triggering receptors. NKp30 is a glycosylated protein and is thought to ;NCR3;NKp30 be selectively expressed in resting and activated natural killer cells. NKp30 is a stimulatory receptor on human NK cells implicated in tumor immunity, and is capable of promoting or terminating dendritic cell maturation. NCR3 may play a role in inflammatory and infectious diseases. Product Information Source Purification Basic Information HEK293 cells > 95% by SDS- PAGE. Description Recombinant Human NCR3/NKp300 Protein is produced by HEK293 cells Endotoxin expression system. The target protein is expressed with sequence (Leu19-Thr138) < 0.1 EU/μg of the protein by LAL of human NCR3/NKp300 (Accession #NP_001138938.1) fused with an Fc, 6×His tag method. at the C-terminus. Formulation Bio-Activity Lyophilized from a 0.22 μm filtered Measured by its binding ability in a functional ELISA. -

Human and Mouse CD Marker Handbook Human and Mouse CD Marker Key Markers - Human Key Markers - Mouse
Welcome to More Choice CD Marker Handbook For more information, please visit: Human bdbiosciences.com/eu/go/humancdmarkers Mouse bdbiosciences.com/eu/go/mousecdmarkers Human and Mouse CD Marker Handbook Human and Mouse CD Marker Key Markers - Human Key Markers - Mouse CD3 CD3 CD (cluster of differentiation) molecules are cell surface markers T Cell CD4 CD4 useful for the identification and characterization of leukocytes. The CD CD8 CD8 nomenclature was developed and is maintained through the HLDA (Human Leukocyte Differentiation Antigens) workshop started in 1982. CD45R/B220 CD19 CD19 The goal is to provide standardization of monoclonal antibodies to B Cell CD20 CD22 (B cell activation marker) human antigens across laboratories. To characterize or “workshop” the antibodies, multiple laboratories carry out blind analyses of antibodies. These results independently validate antibody specificity. CD11c CD11c Dendritic Cell CD123 CD123 While the CD nomenclature has been developed for use with human antigens, it is applied to corresponding mouse antigens as well as antigens from other species. However, the mouse and other species NK Cell CD56 CD335 (NKp46) antibodies are not tested by HLDA. Human CD markers were reviewed by the HLDA. New CD markers Stem Cell/ CD34 CD34 were established at the HLDA9 meeting held in Barcelona in 2010. For Precursor hematopoetic stem cell only hematopoetic stem cell only additional information and CD markers please visit www.hcdm.org. Macrophage/ CD14 CD11b/ Mac-1 Monocyte CD33 Ly-71 (F4/80) CD66b Granulocyte CD66b Gr-1/Ly6G Ly6C CD41 CD41 CD61 (Integrin b3) CD61 Platelet CD9 CD62 CD62P (activated platelets) CD235a CD235a Erythrocyte Ter-119 CD146 MECA-32 CD106 CD146 Endothelial Cell CD31 CD62E (activated endothelial cells) Epithelial Cell CD236 CD326 (EPCAM1) For Research Use Only. -

NK Cell Memory to Cytomegalovirus: Implications for Vaccine Development
Review NK Cell Memory to Cytomegalovirus: Implications for Vaccine Development Calum Forrest 1 , Ariane Gomes 1 , Matthew Reeves 1,* and Victoria Male 2,* 1 Institute of Immunity & Transplantation, UCL, Royal Free Campus, London NW3 2PF, UK; [email protected] (C.F.); [email protected] (A.G.) 2 Department of Metabolism, Digestion and Reproduction, Imperial College London, Chelsea and Westminster Campus, London SW10 9NH, UK * Correspondence: [email protected] (M.R.); [email protected] (V.M.) Received: 24 June 2020; Accepted: 15 July 2020; Published: 20 July 2020 Abstract: Natural killer (NK) cells are innate lymphoid cells that recognize and eliminate virally-infected and cancerous cells. Members of the innate immune system are not usually considered to mediate immune memory, but over the past decade evidence has emerged that NK cells can do this in several contexts. Of these, the best understood and most widely accepted is the response to cytomegaloviruses, with strong evidence for memory to murine cytomegalovirus (MCMV) and several lines of evidence suggesting that the same is likely to be true of human cytomegalovirus (HCMV). The importance of NK cells in the context of HCMV infection is underscored by the armory of NK immune evasion genes encoded by HCMV aimed at subverting the NK cell immune response. As such, ongoing studies that have utilized HCMV to investigate NK cell diversity and function have proven instructive. Here, we discuss our current understanding of NK cell memory to viral infection with a focus on the response to cytomegaloviruses. We will then discuss the implications that this will have for the development of a vaccine against HCMV with particular emphasis on how a strategy that can harness the innate immune system and NK cells could be crucial for the development of a vaccine against this high-priority pathogen. -

Expression Tissue
The Journal of Immunology Killer Cell Ig-Like Receptor and Leukocyte Ig-Like Receptor Transgenic Mice Exhibit Tissue- and Cell-Specific Transgene Expression1 Danny Belkin,* Michaela Torkar,* Chiwen Chang,* Roland Barten,* Mauro Tolaini,† Anja Haude,* Rachel Allen,* Michael J. Wilson,* Dimitris Kioussis,† and John Trowsdale2* To generate an experimental model for exploring the function, expression pattern, and developmental regulation of human Ig-like activating and inhibitory receptors, we have generated transgenic mice using two human genomic clones: 52N12 (a 150-Kb clone encompassing the leukocyte Ig-like receptor (LILR)B1 (ILT2), LILRB4 (ILT3), and LILRA1 (LIR6) genes) and 1060P11 (a 160-Kb clone that contains ten killer cell Ig-like receptor (KIR) genes). Both the KIR and LILR families are encoded within the leukocyte receptor complex, and are involved in immune modulation. We have also produced a novel mAb to LILRA1 to facilitate expression studies. The LILR transgenes were expressed in a similar, but not identical, pattern to that observed in humans: LILRB1 was expressed in B cells, most NK cells, and a small number of T cells; LILRB4 was expressed in a B cell subset; and LILRA1 was found on a ring of cells surrounding B cell areas on spleen sections, consistent with other data showing monocyte/macrophage expression. KIR transgenic mice showed KIR2DL2 expression on a subset of NK cells and T cells, similar to the pattern seen in humans, and expression of KIR2DL4, KIR3DS1, and KIR2DL5 by splenic NK cells. These observations indicate that linked regulatory elements within the genomic clones are sufficient to allow appropriate expression of KIRs in mice, and illustrate that the presence of the natural ligands for these receptors, in the form of human MHC class I proteins, is not necessary for the expression of the KIRs observed in these mice. -

Associated B Cell Lymphoma and Nasopharyngeal Carcinoma
Contribution of tumour cell signalling and the microenvironment to the pathogenesis of EBV- associated B cell lymphoma and nasopharyngeal carcinoma BY MAHA IBRAHIM A thesis submitted to The University of Birmingham For the degree of DOCTOR OF PHILOSOPHY Institute of Cancer and Genomic Sciences College of Medical and Dental Sciences University of Birmingham May 2018 University of Birmingham Research Archive e-theses repository This unpublished thesis/dissertation is copyright of the author and/or third parties. The intellectual property rights of the author or third parties in respect of this work are as defined by The Copyright Designs and Patents Act 1988 or as modified by any successor legislation. Any use made of information contained in this thesis/dissertation must be in accordance with that legislation and must be properly acknowledged. Further distribution or reproduction in any format is prohibited without the permission of the copyright holder. Abstract In this thesis I have explored different components of the pathogenesis of several related EBV associated cancers. In the first part of the thesis I focus on the microenvironment of two of these cancers, nasopharyngeal carcinoma (NPC) and diffuse large B cell lymphoma (DLBCL). Our group has developed a therapeutic vaccine against EBV which has already been shown to be safe in patients with NPC. Therefore, in the first results chapter (chapter 3), I present a description of the phenotyping of expression of the immune microenvironment including immune checkpoint (ICP) genes and MHC class I and class II genes in NPC tissues. I showed for the first time in NPC tissue samples, two types of PD-L1 expressing tumours: diffuse and marginal. -
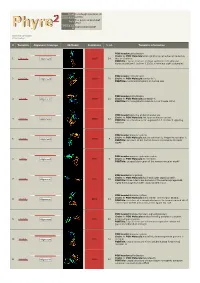
Phyre 2 Results for A1YIY0
Email [email protected] Description A1YIY0 Tue Jul 30 13:19:15 BST Date 2013 Unique Job 1035bc4b501530df ID Detailed template information # Template Alignment Coverage 3D Model Confidence % i.d. Template Information PDB header:cell adhesion Chain: A: PDB Molecule:down syndrome cell adhesion molecule 1 c3dmkA_ Alignment 100.0 14 (dscam) isoform PDBTitle: crystal structure of down syndrome cell adhesion molecule (dscam)2 isoform 1.30.30, n-terminal eight ig domains PDB header:cell adhesion 2 c2om5A_ Alignment 100.0 20 Chain: A: PDB Molecule:contactin 2; PDBTitle: n-terminal fragment of human tax1 PDB header:cell adhesion 3 c3jxaA_ Alignment 100.0 21 Chain: A: PDB Molecule:contactin 4; PDBTitle: immunoglobulin domains 1-4 of mouse cntn4 PDB header:signaling protein/transferase Chain: A: PDB Molecule:tek tyrosine kinase variant; 4 c4k0vA_ 100.0 12 Alignment PDBTitle: structural basis for angiopoietin-1 mediated signaling initiation PDB header:immune system Chain: A: PDB Molecule:natural cytotoxicity triggering receptor 1; 5 c1p6fA_ 100.0 9 Alignment PDBTitle: structure of the human natural cytotoxicity receptor nkp46 PDB header:immune system/receptor 6 c1ollA_ Alignment 99.9 9 Chain: A: PDB Molecule:nk receptor; PDBTitle: extracellular region of the human receptor nkp46 PDB header:viral protein Chain: A: PDB Molecule:hoc head outer capsid protein; 7 c3shsA_ 99.9 18 Alignment PDBTitle: three n-terminal domains of the bacteriophage rb49 highly immunogenic2 outer capsid protein (hoc) PDB header:immune system Chain: E: PDB Molecule:natural -

CD226 T Cells Expressing the Receptors TIGIT and Divergent Phenotypes of Human Regulatory
The Journal of Immunology Divergent Phenotypes of Human Regulatory T Cells Expressing the Receptors TIGIT and CD226 Christopher A. Fuhrman,*,1 Wen-I Yeh,*,1 Howard R. Seay,* Priya Saikumar Lakshmi,* Gaurav Chopra,† Lin Zhang,* Daniel J. Perry,* Stephanie A. McClymont,† Mahesh Yadav,† Maria-Cecilia Lopez,‡ Henry V. Baker,‡ Ying Zhang,x Yizheng Li,{ Maryann Whitley,{ David von Schack,x Mark A. Atkinson,* Jeffrey A. Bluestone,‡ and Todd M. Brusko* Regulatory T cells (Tregs) play a central role in counteracting inflammation and autoimmunity. A more complete understanding of cellular heterogeneity and the potential for lineage plasticity in human Treg subsets may identify markers of disease pathogenesis and facilitate the development of optimized cellular therapeutics. To better elucidate human Treg subsets, we conducted direct transcriptional profiling of CD4+FOXP3+Helios+ thymic-derived Tregs and CD4+FOXP3+Helios2 T cells, followed by comparison with CD4+FOXP32Helios2 T conventional cells. These analyses revealed that the coinhibitory receptor T cell Ig and ITIM domain (TIGIT) was highly expressed on thymic-derived Tregs. TIGIT and the costimulatory factor CD226 bind the common ligand CD155. Thus, we analyzed the cellular distribution and suppressive activity of isolated subsets of CD4+CD25+CD127lo/2 T cells expressing CD226 and/or TIGIT. We observed TIGIT is highly expressed and upregulated on Tregs after activation and in vitro expansion, and is associated with lineage stability and suppressive capacity. Conversely, the CD226+TIGIT2 population was associated with reduced Treg purity and suppressive capacity after expansion, along with a marked increase in IL-10 and effector cytokine production. These studies provide additional markers to delineate functionally distinct Treg subsets that may help direct cellular therapies and provide important phenotypic markers for assessing the role of Tregs in health and disease. -

Single-Cell Profiling Identifies Impaired Adaptive NK Cells Expanded After HCMV Reactivation in Haploidentical HSCT
Single-cell profiling identifies impaired adaptive NK cells expanded after HCMV reactivation in haploidentical HSCT Elisa Zaghi, … , Enrico Lugli, Domenico Mavilio JCI Insight. 2021;6(12):e146973. https://doi.org/10.1172/jci.insight.146973. Research Article Hematology Immunology Graphical abstract Find the latest version: https://jci.me/146973/pdf RESEARCH ARTICLE Single-cell profiling identifies impaired adaptive NK cells expanded after HCMV reactivation in haploidentical HSCT Elisa Zaghi,1 Michela Calvi,1,2 Simone Puccio,3 Gianmarco Spata,1 Sara Terzoli,1 Clelia Peano,4 Alessandra Roberto,3 Federica De Paoli,3 Jasper J.P. van Beek,3 Jacopo Mariotti,5 Chiara De Philippis,5 Barbara Sarina,5 Rossana Mineri,6 Stefania Bramanti,5 Armando Santoro,5 Vu Thuy Khanh Le-Trilling,7 Mirko Trilling,7 Emanuela Marcenaro,8 Luca Castagna,5 Clara Di Vito,1,2 Enrico Lugli,3,9 and Domenico Mavilio1,2 1Unit of Clinical and Experimental Immunology, IRCCS Humanitas Research Hospital, Rozzano, Milan, Italy. 2BIOMETRA, Università degli Studi di Milano, Milan, Italy. 3Laboratory of Translational Immunology, 4Institute of Genetic and Biomedical Research, UoS Milan, National Research Council, and Genomic Unit, 5Bone Marrow Transplant Unit, and 6Molecular Biology Section, Clinical Investigation Laboratory, IRCCS Humanitas Research Hospital, Milan, Italy. 7Institute for Virology, University Hospital Essen, University Duisburg-Essen, Essen, Germany. 8Department of Experimental Medicine, University of Genoa, Genoa, Italy. 9Flow Cytometry Core, IRCCS Humanitas Research Hospital, Milan, Italy. Haploidentical hematopoietic stem cell transplantation (h-HSCT) represents an efficient curative approach for patients affected by hematologic malignancies in which the reduced intensity conditioning induces a state of immunologic tolerance between donor and recipient. -

Natural Killer Cells in Antiviral Immunity
REVIEWS Natural killer cells in antiviral immunity Niklas K. Björkström ✉ , Benedikt Strunz and Hans- Gustaf Ljunggren Abstract | Natural killer (NK) cells play an important role in innate immune responses to viral infections. Here, we review recent insights into the role of NK cells in viral infections, with particular emphasis on human studies. We first discuss NK cells in the context of acute viral infections, with flavivirus and influenza virus infections as examples. Questions related to activation of NK cells, homing to infected tissues and the role of tissue- resident NK cells in acute viral infections are also addressed. Next, we discuss NK cells in the context of chronic viral infections with hepatitis C virus and HIV-1. Also covered is the role of adaptive- like NK cell expansions as well as the appearance of CD56− NK cells in the course of chronic infection. Specific emphasis is then placed in viral infections in patients with primary immunodeficiencies affecting NK cells. Not least, studies in this area have revealed an important role for NK cells in controlling several herpesvirus infections. Finally, we address new data with respect to the activation of NK cells and NK cell function in humans infected with severe acute respiratory syndrome coronavirus 2 (SARS- CoV-2) giving rise to coronavirus disease 2019 (COVID-19). Antibody- dependent Almost 50 years ago, a small number of research groups In humans, mature NK cells are traditionally identified cellular cytotoxicity started to observe unexpected spontaneous cytotoxic as CD3−CD56+ lymphocytes. They were long thought to (ADCC). A mechanism by which activities among lymphocytes. -

Single-Cell RNA Sequencing Demonstrates the Molecular and Cellular Reprogramming of Metastatic Lung Adenocarcinoma
ARTICLE https://doi.org/10.1038/s41467-020-16164-1 OPEN Single-cell RNA sequencing demonstrates the molecular and cellular reprogramming of metastatic lung adenocarcinoma Nayoung Kim 1,2,3,13, Hong Kwan Kim4,13, Kyungjong Lee 5,13, Yourae Hong 1,6, Jong Ho Cho4, Jung Won Choi7, Jung-Il Lee7, Yeon-Lim Suh8,BoMiKu9, Hye Hyeon Eum 1,2,3, Soyean Choi 1, Yoon-La Choi6,10,11, Je-Gun Joung1, Woong-Yang Park 1,2,6, Hyun Ae Jung12, Jong-Mu Sun12, Se-Hoon Lee12, ✉ ✉ Jin Seok Ahn12, Keunchil Park12, Myung-Ju Ahn 12 & Hae-Ock Lee 1,2,3,6 1234567890():,; Advanced metastatic cancer poses utmost clinical challenges and may present molecular and cellular features distinct from an early-stage cancer. Herein, we present single-cell tran- scriptome profiling of metastatic lung adenocarcinoma, the most prevalent histological lung cancer type diagnosed at stage IV in over 40% of all cases. From 208,506 cells populating the normal tissues or early to metastatic stage cancer in 44 patients, we identify a cancer cell subtype deviating from the normal differentiation trajectory and dominating the metastatic stage. In all stages, the stromal and immune cell dynamics reveal ontological and functional changes that create a pro-tumoral and immunosuppressive microenvironment. Normal resident myeloid cell populations are gradually replaced with monocyte-derived macrophages and dendritic cells, along with T-cell exhaustion. This extensive single-cell analysis enhances our understanding of molecular and cellular dynamics in metastatic lung cancer and reveals potential diagnostic and therapeutic targets in cancer-microenvironment interactions. 1 Samsung Genome Institute, Samsung Medical Center, Seoul 06351, Korea. -

Platelets Impair Natural Killer Cell Reactivity and Function in Endometriosis Through Multiple Mechanisms
Human Reproduction, Vol.32, No.4 pp. 794–810, 2017 Advanced Access publication on February 9, 2017 doi:10.1093/humrep/dex014 ORIGINAL ARTICLE Gynaecology Platelets impair natural killer cell reactivity and function in endometriosis through multiple mechanisms Yanbo Du, Xishi Liu, and Sun-Wei Guo* Shanghai Obstetrics and Gynecology Hospital, Fudan University Shanghai College of Medicine, 419 Fangxie Road, Shanghai 200011, China *Correspondence Address. Shanghai Obstetrics and Gynecology Hospital, Fudan University Shanghai College of Medicine, Shanghai 200011, China. Fax: 86-21-6345-5090; E-mail: [email protected] Submitted on October 28, 2016; resubmitted on December 27, 2016; accepted on January 13, 2017 STUDY QUESTION: Do platelets have any role in the reduced cytotoxicity of natural killer (NK) cells in endometriosis? SUMMARY ANSWER: Platelets impair NK cell reactivity and function in endometriosis through multiple mechanisms. WHAT IS KNOWN ALREADY: Platelets play an important role in the development of endometriosis, and platelet-derived transforming growth factor-β1 (TGF-β1) suppresses the expression of NK Group 2, Member D (NKG2D) on NK cells, resulting in reduced cytotoxicity in women with endometriosis. STUDY DESIGN SIZE, DURATION: Experiments on mice with induced endometriosis in which either platelets, NK cells or both were depleted and controls (none depleted). In vitro experiments with NK cells, platelets and, as target cells, endometriotic epithelial cell and endo- metrial stromal cell lines. PARTICIPANTS/MATERIALS SETTING METHODS: Immunohistochemistry analysis of ectopic endometrial tissues from mice with induced endometriosis receiving either platelet depletion (PD), NK cell depletion, or both or none. Immunofluorescence, flow cytometry and gene expression analysis for major histocompatibility complex class I (MHC-I) expression in target cells.