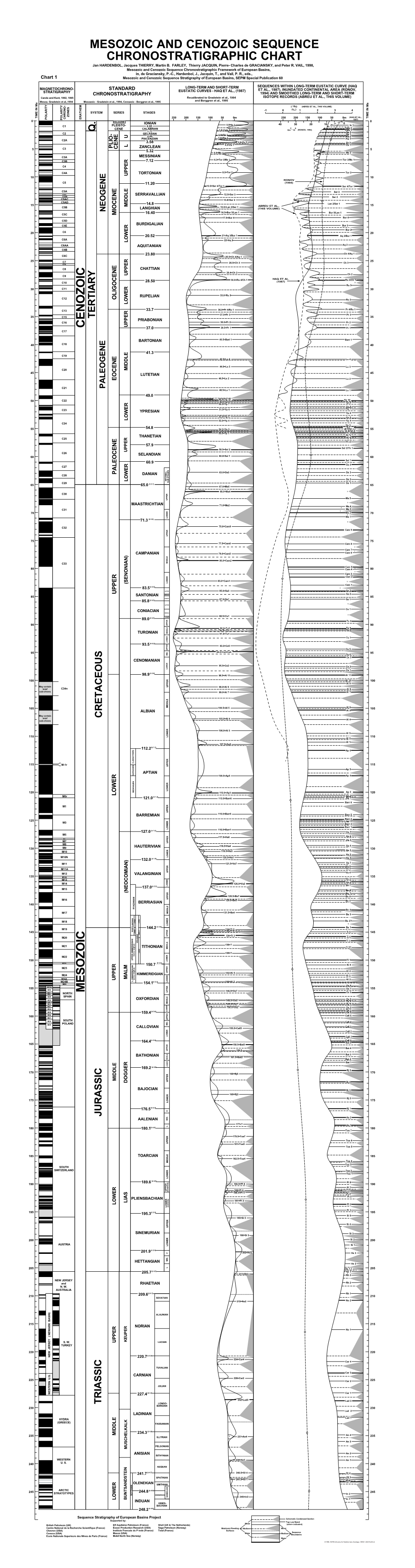
MESOZOIC and CENOZOIC SEQUENCE CHRONOSTRATIGRAPHIC CHART Jan HARDENBOL, Jacques THIERRY, Martin B
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Conodonts and Foraminifers
Journal of Asian Earth Sciences 108 (2015) 117–135 Contents lists available at ScienceDirect Journal of Asian Earth Sciences journal homepage: www.elsevier.com/locate/jseaes An integrated biostratigraphy (conodonts and foraminifers) and chronostratigraphy (paleomagnetic reversals, magnetic susceptibility, elemental chemistry, carbon isotopes and geochronology) for the Permian–Upper Triassic strata of Guandao section, Nanpanjiang Basin, south China ⇑ Daniel J. Lehrmann a, , Leanne Stepchinski a, Demir Altiner b, Michael J. Orchard c, Paul Montgomery d, Paul Enos e, Brooks B. Ellwood f, Samuel A. Bowring g, Jahandar Ramezani g, Hongmei Wang h, Jiayong Wei h, Meiyi Yu i, James D. Griffiths j, Marcello Minzoni k, Ellen K. Schaal l,1, Xiaowei Li l, Katja M. Meyer l,2, Jonathan L. Payne l a Geoscience Department, Trinity University, San Antonio, TX 78212, USA b Department of Geological Engineering, Middle East Technical University, Ankara 06531, Turkey c Natural Resources Canada-Geological Survey of Canada, Vancouver, British Columbia V6B 5J3, Canada d Chevron Upstream Europe, Aberdeen, Scotland, UK e Department of Geology, University of Kansas, Lawrence, KS 66045, USA f Louisiana State University, Baton Rouge, LA 70803, USA g Department of Earth, Atmospheric, and Planetary Sciences, Massachusetts Institute of Technology, Cambridge, MA 02139, USA h Guizhou Geological Survey, Bagongli, Guiyang 550011, Guizhou Province, China i College of Resource and Environment Engineering, Guizhou University, Caijiaguan, Guiyang 550003, Guizhou Province, China j Chemostrat Ltd., 2 Ravenscroft Court, Buttington Cross Enterprise Park, Welshpool, Powys SY21 8SL, UK k Shell International Exploration and Production, 200 N. Dairy Ashford, Houston, TX 77079, USA l Department of Geological and Environmental Sciences, Stanford University, Stanford, CA 94305, USA article info abstract Article history: The chronostratigraphy of Guandao section has served as the foundation for numerous studies of the Received 13 October 2014 end-Permian extinction and biotic recovery in south China. -

Microbialite-Dominated Fossil Associations in Cipit Boulders from Alpe Di Specie and Misurina (St. Cassian Formation, Middle to Upper Triassic, Dolomites, NE Italy)
PUBLICACIÓN CONTINUA ARTÍCULO ORIGINAL © 2019 Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Zaragoza. This is an Open Access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/). TIP Revista Especializada en Ciencias Químico-Biológicas, 22: 1-18, 2019. DOI: 10.22201/fesz.23958723e.2019.0.171 Microbialite-dominated fossil associations in Cipit Boulders from Alpe di Specie and Misurina (St. Cassian Formation, Middle to Upper Triassic, Dolomites, NE Italy) Francisco Sánchez-Beristain1* and Joachim Reitner2 1Museo de Paleontología, Facultad de Ciencias, Universidad Nacional Autónoma de México. Circuito Exterior S/N. Ciudad Universitaria. Coyoacán 04510. Ciudad de México, México. 2Geowissenschaftliches Zentrum der Universität Göttingen, Abt. Geobiologie, Goldschmidtstraße 3. 37077 Göttingen, Germany. E-mail: *[email protected] Abstract In this paper we describe four new fossil associations of “reef” and “reef”-like environments of the St. Cassian Formation (Ladinian-Carnian, Dolomites, NE Italy), based on thirty thin sections from 10 “Cipit boulders” olistoliths, which slided from the Cassian platform into coeval basin sediments. The fossil associations were determined by means of microfacies analysis using point-counting and visual estimation, as well as with aid of statistical methods, based on all fractions with a biotic significance (biomorpha and microbialites). Cluster Analyses in Q-Mode were performed, coupling three algorithms and two indices. In all samples, the main components of the framework are microbialite (average of 75%), and macrofossils (average of 20%), whereas cements and allochtonous components, such as allomicrite, do not represent a significant fraction. Based on both microbialite and fossil content, Chaetetid–microencruster Association, Microbialite–microencruster Association, Dual-type Microbialite Association and Microbialite–Terebella Association, were differentiated. -

Drivers of Beta Diversity in Modern and Ancient Reef-Associated Soft-Bottom Environments
Drivers of beta diversity in modern and ancient reef-associated soft-bottom environments Vanessa Julie Roden1, Martin Zuschin2, Alexander Nützel3,4,5, Imelda M. Hausmann3,4 and Wolfgang Kiessling1 1 GeoZentrum Nordbayern, Section Paleobiology, Friedrich-Alexander University Erlangen-Nürnberg, Erlangen, Germany 2 Department of Palaeontology, University of Vienna, Vienna, Austria 3 SNSB—Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany 4 Department of Earth & Environmental Sciences, Ludwig-Maximilians-Universität München, Munich, Germany 5 GeoBio-Center, Ludwig-Maximilians-Universität München, Munich, Germany ABSTRACT Beta diversity, the compositional variation among communities, is often associated with environmental gradients. Other drivers of beta diversity include stochastic processes, priority effects, predation, or competitive exclusion. Temporal turnover may also explain differences in faunal composition between fossil assemblages. To assess the drivers of beta diversity in reef-associated soft-bottom environments, we investigate community patterns in a Middle to Late Triassic reef basin assemblage from the Cassian Formation in the Dolomites, Northern Italy, and compare results with a Recent reef basin assemblage from the Northern Bay of Safaga, Red Sea, Egypt. We evaluate beta diversity with regard to age, water depth, and spatial distance, and compare the results with a null model to evaluate the stochasticity of these differences. Using pairwise proportional dissimilarity, we find very high beta diversity for the Cassian Formation (0.91 ± 0.02) and slightly lower beta diversity for the Bay of Safaga (0.89 ± 0.04). Null models show that stochasticity only plays a minor role in determining faunal differences. Spatial distance is also irrelevant. Contrary to expectations, there is no tendency of beta Submitted 26 September 2019 Accepted 16 April 2020 diversity to decrease with water depth. -

How to Cite Complete Issue More Information About This Article
TIP. Revista especializada en ciencias químico-biológicas ISSN: 1405-888X ISSN: 2395-8723 Universidad Nacional Autónoma de México, Facultad de Estudios Superiores, Plantel Zaragoza Sánchez-Beristain, Francisco; Reitner, Joachim Microbialite-dominated fossil associations in Cipit Boulders from Alpe di Specie and Misurina (St. Cassian Formation, Middle to Upper Triassic, Dolomites, NE Italy) TIP. Revista especializada en ciencias químico-biológicas, vol. 22, 2019 Universidad Nacional Autónoma de México, Facultad de Estudios Superiores, Plantel Zaragoza DOI: 10.22201/fesz.23958723e.2019.0.171 Available in: http://www.redalyc.org/articulo.oa?id=43265210003 How to cite Complete issue Scientific Information System Redalyc More information about this article Network of Scientific Journals from Latin America and the Caribbean, Spain and Journal's webpage in redalyc.org Portugal Project academic non-profit, developed under the open access initiative PUBLICACIÓN CONTINUA ARTÍCULO ORIGINAL © 2019 Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Zaragoza. This is an Open Access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/). TIP Revista Especializada en Ciencias Químico-Biológicas, 22: 1-18, 2019. DOI: 10.22201/fesz.23958723e.2019.0.171 Microbialite-dominated fossil associations in Cipit Boulders from Alpe di Specie and Misurina (St. Cassian Formation, Middle to Upper Triassic, Dolomites, NE Italy) Francisco Sánchez-Beristain1* and Joachim Reitner2 1Museo de Paleontología, Facultad de Ciencias, Universidad Nacional Autónoma de México. Circuito Exterior S/N. Ciudad Universitaria. Coyoacán 04510. Ciudad de México, México. 2Geowissenschaftliches Zentrum der Universität Göttingen, Abt. Geobiologie, Goldschmidtstraße 3. 37077 Göttingen, Germany. E-mail: *[email protected] Abstract In this paper we describe four new fossil associations of “reef” and “reef”-like environments of the St. -

The Geologic Time Scale Is the Eon
Exploring Geologic Time Poster Illustrated Teacher's Guide #35-1145 Paper #35-1146 Laminated Background Geologic Time Scale Basics The history of the Earth covers a vast expanse of time, so scientists divide it into smaller sections that are associ- ated with particular events that have occurred in the past.The approximate time range of each time span is shown on the poster.The largest time span of the geologic time scale is the eon. It is an indefinitely long period of time that contains at least two eras. Geologic time is divided into two eons.The more ancient eon is called the Precambrian, and the more recent is the Phanerozoic. Each eon is subdivided into smaller spans called eras.The Precambrian eon is divided from most ancient into the Hadean era, Archean era, and Proterozoic era. See Figure 1. Precambrian Eon Proterozoic Era 2500 - 550 million years ago Archaean Era 3800 - 2500 million years ago Hadean Era 4600 - 3800 million years ago Figure 1. Eras of the Precambrian Eon Single-celled and simple multicelled organisms first developed during the Precambrian eon. There are many fos- sils from this time because the sea-dwelling creatures were trapped in sediments and preserved. The Phanerozoic eon is subdivided into three eras – the Paleozoic era, Mesozoic era, and Cenozoic era. An era is often divided into several smaller time spans called periods. For example, the Paleozoic era is divided into the Cambrian, Ordovician, Silurian, Devonian, Carboniferous,and Permian periods. Paleozoic Era Permian Period 300 - 250 million years ago Carboniferous Period 350 - 300 million years ago Devonian Period 400 - 350 million years ago Silurian Period 450 - 400 million years ago Ordovician Period 500 - 450 million years ago Cambrian Period 550 - 500 million years ago Figure 2. -

The Cenozoic Era - Nýlífsöld 65 MY-Present Jarðsaga 2 Ólafur Ingólfsson Origin of the Term: the Tertiary Tertiary System
The Cenozoic Era - Nýlífsöld 65 MY-Present Jarðsaga 2 Ólafur Ingólfsson Origin of the Term: The Tertiary Tertiary System. [1760] Named by Giovanni Arduino Period as the uppermost part of his 65-1.8 MY three-fold subdivision of mountains in northern Italy. The Tertiary became a formal period and system when Lyell published his work describing further subdivisions of the Tertiary. The Tertiary Period is divided into five epochs (tímar): Paleocene (65-56 MY), Eocene (56-34 MY), Oligocene (34-24 MY), Miocene (24-5,3 MY), and Pliocene (5,3-1,8 MY). Confusing set of stratigraphic terms... More than 95% of the Cenozoic era belongs to the Tertiary period. During the 18th century the names Primary, Secondary, and Tertiary were given by Giovanni Arduino to successive rock strata, the Primary being the oldest, the Tertiary the more recent. In 1829 a fourth division, the Quaternary, was added by P. G. Desnoyers. These terms were later abandoned, the Primary becoming the Paleozoic Era, and the Secondary the Mesozoic. But Tertiary and Quaternary were retained for the two main stages of the Cenozoic. Attempts to replace the "Tertiary" with a more reasonable division of “Palaeogene” (early Tertiary) and “Neogene” (later Tertiary and Quaternary) have not been very successful. Stanley uses this division. The World at the K/T Boundary Paleocene plate tectonics During the Paleocene, the inland seas of the Cretaceous Period dry up, exposing large land areas in North America and Eurasia. Australia begins to separate from Antarctica, and Greenland splits from North America. A remnant Tethys Sea persists in the equatorial region. -

Calcareous Nannofossil Zonation and Sequence Stratigraphy of the Jurassic System, Onshore Kuwait
GeoArabia, 2015, v. 20, no. 4, p. 125-180 Gulf PetroLink, Bahrain Calcareous nannofossil zonation and sequence stratigraphy of the Jurassic System, onshore Kuwait Adi P. Kadar, Thomas De Keyser, Nilotpaul Neog and Khalaf A. Karam (with contributions from Yves-Michel Le Nindre and Roger B. Davies) ABSTRACT This paper presents the calcareous nannofossil zonation of the Middle and Upper Jurassic of onshore Kuwait and formalizes current stratigraphic nomenclature. It also interprets the positions of the Jurassic Arabian Plate maximum flooding surfaces (MFS J10 to J110 of Sharland et al., 2001) and sequence boundaries in Kuwait, and correlates them to those in central Saudi Arabia outcrops. This study integrates data from about 400 core samples from 11 wells representing a nearly complete Middle to Upper Jurassic stratigraphic succession. Forty-two nannofossil species were identified using optical microscope techniques. The assemblage contains Tethyan nannofossil markers, which allow application of the Jurassic Tethyan nannofossil biozones. Six zones and five subzones, ranging in age from Middle Aalenian to Kimmeridgian, are established using first and last occurrence events of diagnostic calcareous nannofossil species. A chronostratigraphy of the studied formations is presented, using the revised formal stratigraphic nomenclature. The Marrat Formation is barren of nannofossils. Based on previous studies it is dated as Late Sinemurian–Early Aalenian and contains Middle Toarcian MFS J10. The overlying Dhruma Formation is Middle or Late Aalenian (Zone NJT 8c) or older, to Late Bajocian (Subzone NJT 10a), and contains Lower Bajocian MFS J20. The overlying Sargelu Formation consists of the Late Bajocian (Subzone NJT 10b) Sargelu-Dhruma Transition, and mostly barren Sargelu Limestone in which we place Lower Bathonian MFS J30 near its base. -

Crustacea: Thalassinidea, Brachyura) from Puerto Rico, United States Territory
Bulletin of the Mizunami Fossil Museum, no. 34 (2008), p. 1–15, 6 figs., 1 table. © 2008, Mizunami Fossil Museum New Cretaceous and Cenozoic Decapoda (Crustacea: Thalassinidea, Brachyura) from Puerto Rico, United States Territory Carrie E. Schweitzer1, Jorge Velez-Juarbe2, Michael Martinez3, Angela Collmar Hull1, 4, Rodney M. Feldmann4, and Hernan Santos2 1)Department of Geology, Kent State University Stark Campus, 6000 Frank Ave. NW, North Canton, Ohio, 44720, USA <[email protected]> 2)Department of Geology, University of Puerto Rico, Mayagüez Campus, P. O. Box 9017, Mayagüez, Puerto Rico, 00681 United States Territory <[email protected]> 3)College of Marine Science, University of South Florida, 140 7th Ave. South, St. Petersburg, Florida 33701, USA <[email protected]> 4)Department of Geology, Kent State University, Kent, Ohio 44242, USA <[email protected]> Abstract A large number of recently collected specimens from Puerto Rico has yielded two new species including Palaeoxanthopsis tylotus and Eurytium granulosum, the oldest known occurrence of the latter genus. Cretaceous decapods are reported from Puerto Rico for the first time, and the Cretaceous fauna is similar to that of southern Mexico. Herein is included the first report of Pleistocene decapods from Puerto Rico, which were previously known from other Caribbean localities. The Pleistocene Cardisoma guanhumi is a freshwater crab of the family Gecarcinidae. The freshwater crab families have a poor fossil record; thus, the occurrence is noteworthy and may document dispersal of the crab by humans. Key words: Decapoda, Thalassinidea, Brachyura, Puerto Rico, Cretaceous, Paleogene, Neogene. Introduction than Eocene are not separated by these fault zones and even overlie parts of the fault zones in some areas (Jolly et al., 1998). -

Macroevolutionary Change in the Morphology of the Diatom Fustule Zoe V
This article was downloaded by: [Finkel, Zoe V.] On: 14 September 2010 Access details: Access Details: [subscription number 926896412] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Geomicrobiology Journal Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713722957 Silica Use Through Time: Macroevolutionary Change in the Morphology of the Diatom Fustule Zoe V. Finkela; Benjamin Kotrcb a Environmental Science Program, Mount Allison University, Sackville, New Brunswick, Canada b Department of Earth and Planetary Sciences, Harvard University, Cambridge, Massachusetts, USA Online publication date: 13 September 2010 To cite this Article Finkel, Zoe V. and Kotrc, Benjamin(2010) 'Silica Use Through Time: Macroevolutionary Change in the Morphology of the Diatom Fustule', Geomicrobiology Journal, 27: 6, 596 — 608 To link to this Article: DOI: 10.1080/01490451003702941 URL: http://dx.doi.org/10.1080/01490451003702941 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material. -

Catalogue of Type Fossils in the Geological Museum at the Ohio State University
CATALOGUE OF TYPE FOSSILS IN THE GEOLOGICAL MUSEUM AT THE OHIO STATE UNIVERSITY. HELEN MORNINGSTAR Department of Geology, Ohio State University INTRODUCTION. The following catalogue consists of an alphabetic list of all fossil types (holotypes, paratypes, and cotypes), all subsequently described and figured specimens (plesiotypes), as well as artificial models of type fossils (plastotypes) in the Geological Museum of The Ohio State University. It is patterned directly after the catalogue compiled by Dr. Charles Schuchert, assisted by W. H. Dall, T. W. Stanton, and R. S. Bassler, for the fossil invertebrates in the United States National Museum.1 The catalogue is designed for the convenience of workers in Paleontology and in response to a constant inquiry for informa- tion concerning type specimens of Ohio fossils. Unfortunately, many of the type fossils thought to be located here, according to statements in the older reports of the Geological Survey of Ohio, are missing, and although some of them may have been lost at some former time, it seems probable that many have never been placed in this Museum, but have found their way into other collections. An attempt is now being made to recover as many as possible of the types which have been described in the reports of the Geological Survey of Ohio, and any information concerning tHe location of such specimens will be appreciated. The work of collecting the data for the following catalogue of type fossils extended over a period of several years during which the writer has been engaged in identifying and reclassify- ing the paleontological material in the Geological Museum at The Ohio State University. -

Talexirhynchia, a New Rhynchonellid Genus from the Jurassic Ethiopian Province of Jordan
Talexirhynchia, a new rhynchonellid genus from the Jurassic Ethiopian Province of Jordan Howard R. Feldman, Mena Schemm- Gregory, Mark A. Wilson & Fayez Ahmad Paläontologische Zeitschrift Scientific Contributions to Palaeontology ISSN 0031-0220 Paläontol Z DOI 10.1007/s12542-013-0216-y 1 23 Your article is protected by copyright and all rights are held exclusively by Springer- Verlag Berlin Heidelberg. This e-offprint is for personal use only and shall not be self- archived in electronic repositories. If you wish to self-archive your article, please use the accepted manuscript version for posting on your own website. You may further deposit the accepted manuscript version in any repository, provided it is only made publicly available 12 months after official publication or later and provided acknowledgement is given to the original source of publication and a link is inserted to the published article on Springer's website. The link must be accompanied by the following text: "The final publication is available at link.springer.com”. 1 23 Author's personal copy Pala¨ontol Z DOI 10.1007/s12542-013-0216-y RESEARCH PAPER Talexirhynchia, a new rhynchonellid genus from the Jurassic Ethiopian Province of Jordan Howard R. Feldman • Mena Schemm-Gregory • Mark A. Wilson • Fayez Ahmad Received: 30 May 2013 / Accepted: 28 November 2013 Ó Springer-Verlag Berlin Heidelberg 2013 Abstract A new genus and species of a rhynchonellide scattered on a limy substrate, such as shells and rocks, brachiopod from the Jurassic of Jordan, Talexirhynchia could have served as an attachment site for juveniles. With kadishi gen. et sp. nov., is described. -

Exceptional Fossil Preservation During CO2 Greenhouse Crises? Gregory J
Palaeogeography, Palaeoclimatology, Palaeoecology 307 (2011) 59–74 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Exceptional fossil preservation during CO2 greenhouse crises? Gregory J. Retallack Department of Geological Sciences, University of Oregon, Eugene, Oregon 97403, USA article info abstract Article history: Exceptional fossil preservation may require not only exceptional places, but exceptional times, as demonstrated Received 27 October 2010 here by two distinct types of analysis. First, irregular stratigraphic spacing of horizons yielding articulated Triassic Received in revised form 19 April 2011 fishes and Cambrian trilobites is highly correlated in sequences in different parts of the world, as if there were Accepted 21 April 2011 short temporal intervals of exceptional preservation globally. Second, compilations of ages of well-dated fossil Available online 30 April 2011 localities show spikes of abundance which coincide with stage boundaries, mass extinctions, oceanic anoxic events, carbon isotope anomalies, spikes of high atmospheric carbon dioxide, and transient warm-wet Keywords: Lagerstatten paleoclimates. Exceptional fossil preservation may have been promoted during unusual times, comparable with fi Fossil preservation the present: CO2 greenhouse crises of expanding marine dead zones, oceanic acidi cation, coral bleaching, Trilobite wetland eutrophication, sea level rise, ice-cap melting, and biotic invasions. Fish © 2011 Elsevier B.V. All rights reserved. Carbon dioxide Greenhouse 1. Introduction Zeigler, 1992), sperm (Nishida et al., 2003), nuclei (Gould, 1971)and starch granules (Baxter, 1964). Taphonomic studies of such fossils have Commercial fossil collectors continue to produce beautifully pre- emphasized special places where fossils are exceptionally preserved pared, fully articulated, complex fossils of scientific(Simmons et al., (Martin, 1999; Bottjer et al., 2002).