Neuroprotection by Caffeine and A2A Adenosine Receptor Inactivation in a Model of Parkinson’S Disease
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Neuroprotection by Glial Metabotropic Glutamate Receptors Is Mediated by Transforming Growth Factor-
The Journal of Neuroscience, December 1, 1998, 18(23):9594–9600 Neuroprotection by Glial Metabotropic Glutamate Receptors Is Mediated by Transforming Growth Factor-b V. Bruno,1 G. Battaglia,1 G. Casabona,1 A. Copani,2 F. Caciagli,3 and F. Nicoletti1,2 1Istituto Neurologico Mediterraneo Neuromed, 86077 Pozzilli, Italy, 2Institute of Pharmacology, School of Pharmacy, University of Catania, 95125 Catania, Italy, and 3Department of Pharmacological Sciences, University of Chieti, 66013 Chieti, Italy The medium collected from cultured astrocytes transiently ex- protective activity of DCG-IV or 4C3HPG, as well as the activity posed to the group-II metabotropic glutamate (mGlu) receptor of GM/DCG-IV or GM/4C3HPG; and (3) a transient exposure of 9 9 9 agonists (2S ,1 R ,2 R ,3 R )-2-(2,3-dicarboxycyclopropyl)glycine cultured astrocytes to either DCG-IV or 4C3HPG led to a de- (DCG-IV) or (S)-4-carboxy-3-hydroxyphenylglycine (4C3HPG) layed increase in both intracellular and extracellular levels of is neuroprotective when transferred to mixed cortical cultures TGFb. We therefore conclude that a transient activation of challenged with NMDA (Bruno et al., 1997). The following data group-II mGlu receptors (presumably mGlu3 receptors) in as- indicate that this particular form of neuroprotection is mediated trocytes leads to an increased formation and release of TGFb, by transforming growth factor-b (TGFb). (1) TGFb1 and -b2 which in turn protects neighbor neurons against excitotoxic were highly neuroprotective against NMDA toxicity, and their death. These results offer a new strategy for increasing the local action was less than additive with that produced by the medium production of neuroprotective factors in the CNS. -

Cannabis, the Endocannabinoid System and Immunity—The Journey from the Bedside to the Bench and Back
International Journal of Molecular Sciences Review Cannabis, the Endocannabinoid System and Immunity—The Journey from the Bedside to the Bench and Back Osnat Almogi-Hazan * and Reuven Or Laboratory of Immunotherapy and Bone Marrow Transplantation, Hadassah Medical Center, The Faculty of Medicine, Hebrew University of Jerusalem, Jerusalem 91120, Israel; [email protected] * Correspondence: [email protected] Received: 21 May 2020; Accepted: 19 June 2020; Published: 23 June 2020 Abstract: The Cannabis plant contains numerous components, including cannabinoids and other active molecules. The phyto-cannabinoid activity is mediated by the endocannabinoid system. Cannabinoids affect the nervous system and play significant roles in the regulation of the immune system. While Cannabis is not yet registered as a drug, the potential of cannabinoid-based medicines for the treatment of various conditions has led many countries to authorize their clinical use. However, the data from basic and medical research dedicated to medical Cannabis is currently limited. A variety of pathological conditions involve dysregulation of the immune system. For example, in cancer, immune surveillance and cancer immuno-editing result in immune tolerance. On the other hand, in autoimmune diseases increased immune activity causes tissue damage. Immuno-modulating therapies can regulate the immune system and therefore the immune-regulatory properties of cannabinoids, suggest their use in the therapy of immune related disorders. In this contemporary review, we discuss the roles of the endocannabinoid system in immunity and explore the emerging data about the effects of cannabinoids on the immune response in different pathologies. In addition, we discuss the complexities of using cannabinoid-based treatments in each of these conditions. -

Maresin 1 Promotes Inflammatory Resolution, Neuroprotection, and Functional Neurological Recovery After Spinal Cord Injury
The Journal of Neuroscience, November 29, 2017 • 37(48):11731–11743 • 11731 Development/Plasticity/Repair Maresin 1 Promotes Inflammatory Resolution, Neuroprotection, and Functional Neurological Recovery After Spinal Cord Injury Isaac Francos-Quijorna,1 XEva Santos-Nogueira,1 Karsten Gronert,2 Aaron B. Sullivan,2 XMarcel A. Kopp,3 X Benedikt Brommer,3,4 Samuel David,5 XJan M. Schwab,3,6,7 and XRuben Lo´pez-Vales1 1Departament de Biologia Cel⅐lular, Fisiologia i Immunologia, Institut de Neurociencies, Centro de Investigacio’n Biome’dica en Red sobre Enfermedades Neurodegenerativas (CIBERNED), Universitat Autonoma de Barcelona, 08193 Bellaterra, Catalonia, Spain, 2Vision Science Program, School of Optometry, University of California Berkeley, Berkeley, California 94598, 3Spinal Cord Alliance Berlin (SCAB) and Department of Neurology and Experimental Neurology, Charite´ Campus Mitte, Clinical and Experimental Spinal Cord Injury Research Laboratory (Neuroparaplegiology), Charite´-Universita¨tsmedizin Berlin, 10117 Berlin, Germany, 4F.M. Kirby Neurobiology Center, Boston Children’s Hospital, and Department of Neurology, Harvard Medical School, Boston, Massachusetts 02115, 5Centre for Research in Neuroscience, The Research Institute of the McGill University Health Centre, H3G1A4 Montreal, Canada, and 6Department of Neurology, Spinal Cord Injury Division, and 7Department of Neuroscience and Center for Brain and Spinal Cord Repair, Department of Physical Medicine and Rehabilitation, The Neurological Institute, The Ohio State University, Wexner Medical Centre, Columbus, Ohio 43210 Resolution of inflammation is defective after spinal cord injury (SCI), which impairs tissue integrity and remodeling and leads to functional deficits. Effective pharmacological treatments for SCI are not currently available. Maresin 1 (MaR1) is a highly conserved specialized proresolving mediator (SPM) hosting potent anti-inflammatory and proresolving properties with potent tissue regenerative actions. -

Advances in Non-Dopaminergic Treatments for Parkinson's Disease
REVIEW ARTICLE published: 22 May 2014 doi: 10.3389/fnins.2014.00113 Advances in non-dopaminergic treatments for Parkinson’s disease Sandy Stayte 1,2 and Bryce Vissel 1,2* 1 Neuroscience Department, Neurodegenerative Disorders Laboratory, Garvan Institute of Medical Research, Sydney, NSW, Australia 2 Faculty of Medicine, University of New South Wales, Sydney, NSW, Australia Edited by: Since the 1960’s treatments for Parkinson’s disease (PD) have traditionally been directed Eero Vasar, University of Tartu, to restore or replace dopamine, with L-Dopa being the gold standard. However, chronic Estonia L-Dopa use is associated with debilitating dyskinesias, limiting its effectiveness. This has Reviewed by: resulted in extensive efforts to develop new therapies that work in ways other than Andrew Harkin, Trinity College Dublin, Ireland restoring or replacing dopamine. Here we describe newly emerging non-dopaminergic Sulev Kõks, University of Tartu, therapeutic strategies for PD, including drugs targeting adenosine, glutamate, adrenergic, Estonia and serotonin receptors, as well as GLP-1 agonists, calcium channel blockers, iron Pille Taba, Universoty of Tartu, chelators, anti-inflammatories, neurotrophic factors, and gene therapies. We provide a Estonia Pekka T. Männistö, University of detailed account of their success in animal models and their translation to human clinical Helsinki, Finland trials. We then consider how advances in understanding the mechanisms of PD, genetics, *Correspondence: the possibility that PD may consist of multiple disease states, understanding of the Bryce Vissel, Neuroscience etiology of PD in non-dopaminergic regions as well as advances in clinical trial design Department, Neurodegenerative will be essential for ongoing advances. We conclude that despite the challenges ahead, Disorders Laboratory, Garvan Institute of Medical Research, patients have much cause for optimism that novel therapeutics that offer better disease 384 Victoria Street, Darlinghurst, management and/or which slow disease progression are inevitable. -

Caffeine Induces Neurobehavioral Effects Through Modulating Neurotransmitters
Saudi Pharmaceutical Journal 28 (2020) 445–451 Contents lists available at ScienceDirect Saudi Pharmaceutical Journal journal homepage: www.sciencedirect.com Review Caffeine induces neurobehavioral effects through modulating neurotransmitters Fawaz Alasmari Department of Pharmacology and Toxicology, College of Pharmacy, King Saud University, Riyadh 11451, Saudi Arabia article info abstract Article history: Evidence demonstrates that chronic caffeine exposure, primarily through consumption of coffee or tea, Received 19 November 2019 leads to increased alertness and anxiety. Preclinical and clinical studies showed that caffeine induced Accepted 12 February 2020 beneficial effects on mood and cognition. Other studies using molecular techniques have reported that Available online 17 February 2020 caffeine exhibited neuroprotective effects in animal models by protecting dopaminergic neurons. Moreover, caffeine interacts with dopaminergic system, which leads to improvements in neurobehavioral Keywords: measures in animal models of depression or attention deficit hyperactivity disorder (ADHD). Caffeine Glutamatergic receptors have been found to be involved on the neurobiological effects of caffeine. Neurobehavioral effects Additionally, caffeine has been found to suppress the inhibitory (GABAergic) activity and modulate Neurotransmitters Dopamine GABA receptors. Studies have also found that modulating these neurotransmitters leads to neurobehav- Glutamate ioral effects. The linkage between the modulatory role of caffeine on neurotransmitters and neurobehav- GABA ioral effects has not been fully discussed. The purpose of this review is to discuss in detail the role of neurotransmitters in the effects of caffeine on neurobehavioral disorders. Ó 2020 The Author. Published by Elsevier B.V. on behalf of King Saud University. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/). -
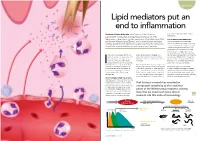
Lipid Mediators Put an End to Inflammation
Biochemistry Lipid mediators put an end to inflammation is the Director of the Center for than merely antagonising inflammatory Professor Charles N Serhan mechanisms. Experimental Therapeutics and Reperfusion Injury (CETRI) in the Department of Anesthesiology, Perioperative and Pain Medicine at BWH ACTIVE RESOLUTION MEDIATORS and Harvard Medical School, Boston. In his current research he leads a Prof Serhan’s research has discovered multidisciplinary team of experts whose goal is to reveal the mechanisms potent anti-inflammatory and pro-resolving of resolution of acute inflammation and tissue injury in the body. compounds made by the body that he calls specialised pro-resolving mediators (SPM). One group of SPM comes from the lipids that that we eat – namely from essential omega-3 fatty acids. Within this group, we nflammation is an essential part of our effective treatments for diseases with can distinguish between resolvins, protectins defence against infections, cancer and aberrant on-going inflammation such as and maresins. SPMs have been measured other attacks to our body’s normal Alzheimer's disease, cardiovascular disease at bioactive levels in body fluids like human function. Ideally, any inflammatory or arthritis. tears and blood, as well as tissues like the process should be self-limiting so that brain, lymph nodes, spleen and fat. the body returns to its previous healthy For a long time, there was only a vague idea Istate. But how does this happen? Prof about the passive mechanisms that stop the Interestingly, aspirin can enhance the Serhan's research aims to elucidate the inflammatory response. This has changed in formation of specific SPM: aspirin-triggered mechanisms underlying the resolution recent years thanks to researchers like Prof resolvins and protectins have been described. -

The Interaction of Selective A1 and A2A Adenosine Receptor Antagonists with Magnesium and Zinc Ions in Mice: Behavioural, Biochemical and Molecular Studies
International Journal of Molecular Sciences Article The Interaction of Selective A1 and A2A Adenosine Receptor Antagonists with Magnesium and Zinc Ions in Mice: Behavioural, Biochemical and Molecular Studies Aleksandra Szopa 1,* , Karolina Bogatko 1, Mariola Herbet 2 , Anna Serefko 1 , Marta Ostrowska 2 , Sylwia Wo´sko 1, Katarzyna Swi´ ˛ader 3, Bernadeta Szewczyk 4, Aleksandra Wla´z 5, Piotr Skałecki 6, Andrzej Wróbel 7 , Sławomir Mandziuk 8, Aleksandra Pochodyła 3, Anna Kudela 2, Jarosław Dudka 2, Maria Radziwo ´n-Zaleska 9, Piotr Wla´z 10 and Ewa Poleszak 1,* 1 Chair and Department of Applied and Social Pharmacy, Laboratory of Preclinical Testing, Medical University of Lublin, 1 Chod´zkiStreet, PL 20–093 Lublin, Poland; [email protected] (K.B.); [email protected] (A.S.); [email protected] (S.W.) 2 Chair and Department of Toxicology, Medical University of Lublin, 8 Chod´zkiStreet, PL 20–093 Lublin, Poland; [email protected] (M.H.); [email protected] (M.O.); [email protected] (A.K.) [email protected] (J.D.) 3 Chair and Department of Applied and Social Pharmacy, Medical University of Lublin, 1 Chod´zkiStreet, PL 20–093 Lublin, Poland; [email protected] (K.S.);´ [email protected] (A.P.) 4 Department of Neurobiology, Polish Academy of Sciences, Maj Institute of Pharmacology, 12 Sm˛etnaStreet, PL 31–343 Kraków, Poland; [email protected] 5 Department of Pathophysiology, Medical University of Lublin, 8 Jaczewskiego Street, PL 20–090 Lublin, Poland; [email protected] Citation: Szopa, A.; Bogatko, K.; 6 Department of Commodity Science and Processing of Raw Animal Materials, University of Life Sciences, Herbet, M.; Serefko, A.; Ostrowska, 13 Akademicka Street, PL 20–950 Lublin, Poland; [email protected] M.; Wo´sko,S.; Swi´ ˛ader, K.; Szewczyk, 7 Second Department of Gynecology, 8 Jaczewskiego Street, PL 20–090 Lublin, Poland; B.; Wla´z,A.; Skałecki, P.; et al. -

A Selective Trkb Agonist with Potent Neurotrophic Activities by 7,8-Dihydroxyflavone
A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone Sung-Wuk Janga, Xia Liua, Manuel Yepesb, Kennie R. Shepherdc, Gary W. Millerc, Yang Liud, W. David Wilsond, Ge Xiaoe, Bruno Blanchif, Yi E. Sunf, and Keqiang Yea,1 aDepartment of Pathology and Laboratory Medicine and bDepartment of Neurology, Emory University School of Medicine, Atlanta, GA 30322; cDepartment of Environmental and Occupational Health, Rollins Public School of Health, Emory University, Atlanta, GA 30322; dDepartment of Chemistry, Georgia State University, Atlanta, GA 30302; eCenters for Disease Control and Prevention, Atlanta, GA 30322; and fNeuropsychiatric Institute, Medical Retardation Research Center, University of California, Los Angeles, Los Angeles, CA 90095 Edited* by Solomon Snyder, Johns Hopkins University School of Medicine, Baltimore, MD, and approved December 30, 2009 (received for review November 25, 2009) Brain-derived neurotrophic factor (BDNF), a cognate ligand for the Y490 phosphorylation and Akt activation in both T48 and T62 tyrosine kinase receptor B (TrkB) receptor, mediates neuronal sur- cell lines, whereas only faint Trk activation and no Akt phos- vival, differentiation, synaptic plasticity, and neurogenesis. However, phorylation were demonstrated in T17 clones that stably express BDNF has a poor pharmacokinetic profile that limits its therapeutic TrkA. As expected, BDNF substantially decreased apoptosis in potential. Here we report the identification of 7,8-dihydroxyflavone T48 cells compared with the parental SN56 cells. Even in the as a bioactive high-affinity TrkB agonist that provokes receptor absence of BDNF, T48 cells were slightly resistant to apoptosis, dimerization and autophosphorylation and activation of down- indicating that overexpression of TrkB weakly suppresses cas- stream signaling. -

Mglur5, CB1 and Neuroprotection
www.impactjournals.com/oncotarget/ Oncotarget, 2017, Vol. 8, (No. 3), pp: 3768-3769 Editorial: Neuroscience mGluR5, CB1 and neuroprotection Toniana G. Carvalho, Juliana G. Doria and Fabiola M. Ribeiro The metabotropic glutamate receptor 5 (mGluR5) is either mGluR5-/- or CB1 knockdown neurons. In addition, a Gαq/11-coupled receptor, mainly found at the postsynaptic neuroprotection by CDPPB, URB597 and JZL184 was site. mGluR5 stimulation leads to the activation of dependent on the activation of pathways that lead to the phospholipase Cβ1 (PLCβ), promoting diacylglicerol stimulation of AKT and ERK1/2, but was independent of 2+ (DAG) and inositol 1,4,5-trisphosphate (IP3) formation, alterations in intracellular Ca concentration or glutamate which leads to the release of Ca2+ from the intracellular release. stores and the activation of protein kinases, including In several neurodegenerative diseases, neuronal protein kinase C. Additionally, stimulation of mGluR5 also cell death is preceded by synaptic loss, which is usually triggers the activation of other cell signaling pathways that responsible for the early cognitive deficits. The study by are important for cell proliferation and survival, such as Batista et al, 2016 [6], showed that CDPPB protected the the activation of the extracellular signal regulated protein postsynaptic site and that this protection was blocked kinase (ERK) and AKT. Recently, we have demonstrated by MPEP and AM251. In addition, JZL184 protected that the mGluR5 positive allosteric modulator (PAM), the presynaptic terminals and, to a lesser extent, the CDPPB, activates AKT without increasing intracellular postsynaptic site. Curiously, AM251 reversed JZL- Ca2+ and protects neurons from glutamate-induced mediated protection of both the pre and postsynapse, while neuronal cell death [1]. -

A Differential Role for the Adenosine A2A Receptor in Opiate Reinforcement Vs Opiate-Seeking Behavior
Neuropsychopharmacology (2009) 34, 844–856 & 2009 Nature Publishing Group All rights reserved 0893-133X/09 $32.00 www.neuropsychopharmacology.org A Differential Role for the Adenosine A2A Receptor in Opiate Reinforcement vs Opiate-Seeking Behavior 1,2 2 1,3 4 Robyn Mary Brown , Jennifer Lynn Short , Michael Scott Cowen , Catherine Ledent and ,1,3 Andrew John Lawrence* 1Brain Injury and Repair Group, Howard Florey Institute, University of Melbourne, Parkville, VIC, Australia; 2Monash Institute of Pharmaceutical 3 4 Sciences, Parkville, VIC, Australia; Centre for Neuroscience, University of Melbourne, Parkville, VIC, Australia; Faculte de Medecine, Institut de Recherche Interdisciplinaire, Universite de Bruxelles, Brussels, Belgium The adenosine A2A receptor is specifically enriched in the medium spiny neurons that make up the ‘indirect’ output pathway from the ventral striatum, a structure known to have a crucial, integrative role in processes such as reward, motivation, and drug-seeking behavior. In the present study we investigated the impact of adenosine A receptor deletion on behavioral responses to morphine in a number of 2A reward-related paradigms. The acute, rewarding effects of morphine were evaluated using the conditioned place preference paradigm. Operant self-administration of morphine on both fixed and progressive ratio schedules as well as cue-induced drug-seeking was assessed. In addition, the acute locomotor response to morphine as well as sensitization to morphine was evaluated. Decreased morphine self- administration and breakpoint in A2A knockout mice was observed. These data support a decrease in motivation to consume the drug, perhaps reflecting diminished rewarding effects of morphine in A2A knockout mice. In support of this finding, a place preference to morphine was not observed in A knockout mice but was present in wild-type mice. -

Therapeutic Effects of Specialized Pro-Resolving Lipids Mediators On
antioxidants Review Therapeutic Effects of Specialized Pro-Resolving Lipids Mediators on Cardiac Fibrosis via NRF2 Activation 1, 1,2, 2, Gyeoung Jin Kang y, Eun Ji Kim y and Chang Hoon Lee * 1 Lillehei Heart Institute, University of Minnesota, Minneapolis, MN 55455, USA; [email protected] (G.J.K.); [email protected] (E.J.K.) 2 College of Pharmacy, Dongguk University, Seoul 04620, Korea * Correspondence: [email protected]; Tel.: +82-31-961-5213 Equally contributed. y Received: 11 November 2020; Accepted: 9 December 2020; Published: 10 December 2020 Abstract: Heart disease is the number one mortality disease in the world. In particular, cardiac fibrosis is considered as a major factor causing myocardial infarction and heart failure. In particular, oxidative stress is a major cause of heart fibrosis. In order to control such oxidative stress, the importance of nuclear factor erythropoietin 2 related factor 2 (NRF2) has recently been highlighted. In this review, we will discuss the activation of NRF2 by docosahexanoic acid (DHA), eicosapentaenoic acid (EPA), and the specialized pro-resolving lipid mediators (SPMs) derived from polyunsaturated lipids, including DHA and EPA. Additionally, we will discuss their effects on cardiac fibrosis via NRF2 activation. Keywords: cardiac fibrosis; NRF2; lipoxins; resolvins; maresins; neuroprotectins 1. Introduction Cardiovascular disease is the leading cause of death worldwide [1]. Cardiac fibrosis is a major factor leading to the progression of myocardial infarction and heart failure [2]. Cardiac fibrosis is characterized by the net accumulation of extracellular matrix proteins in the cardiac stroma and ultimately impairs cardiac function [3]. Therefore, interest in substances with cardioprotective activity continues. -

Neuroprotection by Δ9-Tetrahydrocannabinol, the Main Active Compound in Marijuana, Against Ouabain-Induced in Vivo Excitotoxici
The Journal of Neuroscience, September 1, 2001, 21(17):6475–6479 Neuroprotection by ⌬9-Tetrahydrocannabinol, the Main Active Compound in Marijuana, against Ouabain-Induced In Vivo Excitotoxicity M. van der Stelt,1 W. B. Veldhuis,2,3 P. R. Ba¨r,3 G. A. Veldink,1 J. F. G. Vliegenthart,1 and K. Nicolay2 1Department of Bio-Organic Chemistry, Bijvoet Center for Biomolecular Research, 3584 CH, Utrecht University, Utrecht, The Netherlands, 2Department of Experimental In Vivo NMR, Image Sciences Institute, 3584 CJ, Utrecht, University Medical Center Utrecht, The Netherlands, and 3Department of Experimental Neurology, University Medical Center Utrecht, 3584 CX, Utrecht, The Netherlands Excitotoxicity is a paradigm used to explain the biochemical SR141716 prevented the neuroprotective actions of ⌬9-THC, events in both acute neuronal damage and in slowly progres- indicating that ⌬9-THC afforded protection to neurons via the ⌬9 sive, neurodegenerative diseases. Here, we show in a longitu- CB1 receptor. In -THC-treated rats the volume of astrogliotic ⌬9 dinal magnetic resonance imaging study that -tetrahydro- tissue was 36% smaller. The CB1 receptor antagonist did not cannabinol (⌬9-THC), the main active compound in marijuana, block this effect. These results provide evidence that the can- reduces neuronal injury in neonatal rats injected intracerebrally nabinoid system can serve to protect the brain against with the Na ϩ/K ϩ-ATPase inhibitor ouabain to elicit excitotox- neurodegeneration. icity. In the acute phase ⌬9-THC reduced the volume of cyto- toxic edema by 22%. After 7 d, 36% less neuronal damage was Key words: anandamide; astrogliosis; cannabinoid; excitotox- observed in treated rats compared with control animals.