ANH Vxx 07 Lowres.D15d742.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Proceedings of the Biological Society of Washington
PROC. BIOL. SOC. WASH. 103(1), 1990, pp. 248-253 SIX NEW COMBINATIONS IN BACCHAROIDES MOENCH AND CYANTHILLIUM "SUJME (VERNONIEAE: ASTERACEAE) Harold Robinson Abstract.— ThvQQ species, Vernonia adoensis Schultz-Bip. ex Walp., V. gui- neensis Benth., and V. lasiopus O. HofFm. in Engl., are transferred to the genus Baccharoides Moench, and three species, Conyza cinerea L., C. patula Ait., and Herderia stellulifera Benth. are transferred to the genus Cyanthillium Blume. The present paper provides six new com- tinct from the Western Hemisphere mem- binations of Old World Vemonieae that are bers of that genus. Although generic limits known to belong to the genera Baccharoides were not discussed by Jones, his study placed Moench and Cyanthillium Blume. The ap- the Old World Vernonia in a group on the plicability of these generic names to these opposite side the basic division in the genus species groups was first noted by the author from typical Vernonia in the eastern United almost ten years ago (Robinson et al. 1 980), States. Subsequent studies by Jones (1979b, and it was anticipated that other workers 1981) showed that certain pollen types also more familiar with the paleotropical mem- were restricted to Old World members of bers of the Vernonieae would provide the Vernonia s.l., types that are shared by some necessary combinations. A recent study of Old World members of the tribe tradition- eastern African members of the tribe by Jef- ally placed in other genera. The characters frey (1988) also cites these generic names as noted by Jones have been treated by the synonyms under his Vernonia Group 2 present author as evidence of a basic divi- subgroup C and Vernonia Group 4, al- sion in the Vernonieae between groups that though he retains the broad concept of Ver- have included many genera in each hemi- nonia. -

Brachychiton Discolor Lacebark
Plant of the Week Brachychiton discolor Lacebark In late spring and early summer, Sydney is blessed with flowering trees: mauve blue Jacarandas, scarlet Illawarra Flame Trees and golden Silky Oaks. The Lacebark (Brachychiton discolor) also flowers at this time of year but somehow we often miss the lovely dusky pink and brown flowers of this most elegant of Australian trees. The Lacebark is closely related to the Illawarra Flame (Brachychiton acerifolius), Kurrajong (B. populneus) and the Queensland Bottle Tree (B. rupestris), all of which are now included in the plant family Malvaceae although until recently, were included in the Sterculiaceae, a family of tropical and subtropical distribution. Lacebarks can be found in rainforests along the coast and ranges of eastern Australia, from Paterson in northern NSW, to Mackay in North Queensland. There is also an isolated community on Cape York Peninsula1. They are very popular as garden plants as they don’t grow too tall, can cope with a wide range of soils and can survive hot and dry conditions. They drop their leaves just once a year prior to flowering and it is then that you notice the striking contrast between their straight round trunks and unusual branching architecture which is so very different from that of eucalypts with which we are more familiar. 1Wikipedia: http://en.wikipedia.org/wiki/Brachychiton_discolor Map modified from Australia’s Virtual Herbarium: http://www.chah.gov.au/avh/avhServlet Text and photographs: Alison Downing & Kevin Downing, 19.11.2011 Downing Herbarium, Department of Biological Sciences . -

Winter Edition 2020 - 3 in This Issue: Office Bearers for 2017
1 Australian Plants Society Armidale & District Group PO Box 735 Armidale NSW 2350 web: www.austplants.com.au/Armidale e-mail: [email protected] Crowea exalata ssp magnifolia image by Maria Hitchcock Winter Edition 2020 - 3 In this issue: Office bearers for 2017 ......p1 Editorial …...p2Error! Bookmark not defined. New Website Arrangements .…..p3 Solstice Gathering ......p4 Passion, Boers & Hibiscus ......p5 Wollomombi Falls Lookout ......p7 Hard Yakka ......p8 Torrington & Gibraltar after fires ......p9 Small Eucalypts ......p12 Drought tolerance of plants ......p15 Armidale & District Group PO Box 735, Armidale NSW 2350 President: Vacant Vice President: Colin Wilson Secretary: Penelope Sinclair Ph. 6771 5639 [email protected] Treasurer: Phil Rose Ph. 6775 3767 [email protected] Membership: Phil Rose [email protected] 2 Markets in the Mall, Outings, OHS & Environmental Officer and Arboretum Coordinator: Patrick Laher Ph: 0427327719 [email protected] Newsletter Editor: John Nevin Ph: 6775218 [email protected],net.au Meet and Greet: Lee Horsley Ph: 0421381157 [email protected] Afternoon tea: Deidre Waters Ph: 67753754 [email protected] Web Master: Eric Sinclair Our website: http://www.austplants.com.au From the Editor: We have certainly had a memorable year - the worst drought in living memory followed by the most extensive bushfires seen in Australia, and to top it off, the biggest pandemic the world has seen in 100 years. The pandemic has made essential self distancing and quarantining to arrest the spread of the Corona virus. As a result, most APS activities have been shelved for the time being. Being in isolation at home has been a mixed blessing. -

The Geranium Family, Geraniaceae, and the Mallow Family, Malvaceae
THE GERANIUM FAMILY, GERANIACEAE, AND THE MALLOW FAMILY, MALVACEAE TWO SOMETIMES CONFUSED FAMILIES PROMINENT IN SOME MEDITERRANEAN CLIMATE AREAS The Geraniaceae is a family of herbaceous plants or small shrubs, sometimes with succulent stems • The family is noted for its often palmately veined and lobed leaves, although some also have pinnately divided leaves • The leaves all have pairs of stipules at their base • The flowers may be regular and symmetrical or somewhat irregular • The floral plan is 5 separate sepals and petals, 5 or 10 stamens, and a superior ovary • The most distinctive feature is the beak of fused styles on top of the ovary Here you see a typical geranium flower This nonnative weedy geranium shows the styles forming a beak The geranium family is also noted for its seed dispersal • The styles either actively eject the seeds from each compartment of the ovary or… • They twist and embed themselves in clothing and fur to hitch a ride • The Geraniaceae is prominent in the Mediterranean Basin and the Cape Province of South Africa • It is also found in California but few species here are drought tolerant • California does have several introduced weedy members Here you see a geranium flinging the seeds from sections of the ovary when the styles curl up Three genera typify the Geraniaceae: Erodium, Geranium, and Pelargonium • Erodiums (common name filaree or clocks) typically have pinnately veined, sometimes dissected leaves; many species are weeds in California • Geraniums (that is, the true geraniums) typically have palmately veined leaves and perfectly symmetrical flowers. Most are herbaceous annuals or perennials • Pelargoniums (the so-called garden geraniums or storksbills) have asymmetrical flowers and range from perennials to succulents to shrubs The weedy filaree, Erodium cicutarium, produces small pink-purple flowers in California’s spring grasslands Here are the beaked unripe fruits of filaree Many of the perennial erodiums from the Mediterranean make well-behaved ground covers for California gardens Here are the flowers of the charming E. -

TML Propagation Protocols
PROPAGATION PROTOCOLS This document is intended as a guide for Tamborine Mountain Landcare members who wish to assist our regeneration projects by growing some of the plants needed. It is a work in progress so if you have anything to add to the protocols – for example a different but successful way of propagating and growing a particular plant – then please give it to Julie Lake so she can add it to the document. The idea is that our shared knowledge and experience can become a valuable part of TML's intellectual property as well as a useful source of knowledge for members. As there are many hundreds of plants native to Tamborine Mountain, the protocols list will take a long time to complete, with growing information for each plant added alphabetically as time permits. While the list is being compiled by those members with competence in this field, any TML member with a query about propagating a particular plant can post it on the website for other me mb e r s to answer. To date, only protocols for trees and shrubs have been compiled. Vines and ferns will be added later. Fruiting times given are usual for the species but many rainforest plants flower and fruit opportunistically, according to weather and other conditions unknown to us, thus fruit can be produced at any time of year. Finally, if anyone would like a copy of the protocols, contact Julie on [email protected] and she’ll send you one. ………………….. Growing from seed This is the best method for most plants destined for regeneration projects for it is usually fast, easy and ensures genetic diversity in the regenerated landscape. -

Plant Mobility in the Mesozoic Disseminule Dispersal Strategies Of
Palaeogeography, Palaeoclimatology, Palaeoecology 515 (2019) 47–69 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Plant mobility in the Mesozoic: Disseminule dispersal strategies of Chinese and Australian Middle Jurassic to Early Cretaceous plants T ⁎ Stephen McLoughlina, , Christian Potta,b a Palaeobiology Department, Swedish Museum of Natural History, Box 50007, 104 05 Stockholm, Sweden b LWL - Museum für Naturkunde, Westfälisches Landesmuseum mit Planetarium, Sentruper Straße 285, D-48161 Münster, Germany ARTICLE INFO ABSTRACT Keywords: Four upper Middle Jurassic to Lower Cretaceous lacustrine Lagerstätten in China and Australia (the Daohugou, Seed dispersal Talbragar, Jehol, and Koonwarra biotas) offer glimpses into the representation of plant disseminule strategies Zoochory during that phase of Earth history in which flowering plants, birds, mammals, and modern insect faunas began to Anemochory diversify. No seed or foliage species is shared between the Northern and Southern Hemisphere fossil sites and Hydrochory only a few species are shared between the Jurassic and Cretaceous assemblages in the respective regions. Free- Angiosperms sporing plants, including a broad range of bryophytes, are major components of the studied assemblages and Conifers attest to similar moist growth habitats adjacent to all four preservational sites. Both simple unadorned seeds and winged seeds constitute significant proportions of the disseminule diversity in each assemblage. Anemochory, evidenced by the development of seed wings or a pappus, remained a key seed dispersal strategy through the studied interval. Despite the rise of feathered birds and fur-covered mammals, evidence for epizoochory is minimal in the studied assemblages. Those Early Cretaceous seeds or detached reproductive structures bearing spines were probably adapted for anchoring to aquatic debris or to soft lacustrine substrates. -

Argemone Mexicana L.)
Electronic Journal of Biotechnology ISSN: 0717-3458 Vol.11 No.1, Issue of January 15, 2008 © 2008 by Pontificia Universidad Católica de Valparaíso -- Chile Received March 21, 2007 / Accepted Mayo 20, 2007 DOI: 10.2225/vol11-issue1-fulltext-3 SHORT COMMUNICATION Agrobacterium-mediated transient transformation of Mexican prickly poppy (Argemone mexicana L.) Gregorio Godoy-Hernández* Unidad de Bioquímica y Biología Molecular de Plantas Centro de Investigación Científica de Yucatán A.C. C. 43 No. 130, Chuburná de Hidalgo 97200 Mérida, Yucatán, México Tel: 999 9428330 Fax: 999 9 81 39 00 E-mail: [email protected] Elidé Avilés-Berzunza Unidad de Bioquímica y Biología Molecular de Plantas Centro de Investigación Científica de Yucatán A.C. C. 43 No. 130, Chuburná de Hidalgo 97200 Mérida, Yucatán, México Tel: 999 9428330 Fax: 999 9 81 39 00 E-mail: [email protected] Mildred Carrillo-Pech Unidad de Bioquímica y Biología Molecular de Plantas Centro de Investigación Científica de Yucatán A.C. C. 43 No. 130, Chuburná de Hidalgo 97200 Mérida, Yucatán, México Tel: 999 9428330 Fax: 999 9 81 39 00 E-mail: [email protected] Felipe Vázquez-Flota Unidad de Bioquímica y Biología Molecular de Plantas Centro de Investigación Científica de Yucatán A.C. C. 43 No. 130, Chuburná de Hidalgo 97200 Mérida, Yucatán, México Tel: 999 9428330 Fax: 999 9 81 39 00 E-mail: [email protected] Website: http://www.cicy.mx Financial support: Consejo Nacional de Ciencia y Tecnología (CONACYT) from México (Grant No. 28643-B). Keywords: Argemone mexicana, β-glucuronidase, benzylisoquinoline alkaloids, genetic transformation, neomycin phosphotransferase II. -

Vascular Plant and Vertebrate Inventory of Chiricahua National Monument
In Cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Chiricahua National Monument Open-File Report 2008-1023 U.S. Department of the Interior U.S. Geological Survey National Park Service This page left intentionally blank. In cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Chiricahua National Monument By Brian F. Powell, Cecilia A. Schmidt, William L. Halvorson, and Pamela Anning Open-File Report 2008-1023 U.S. Geological Survey Southwest Biological Science Center Sonoran Desert Research Station University of Arizona U.S. Department of the Interior School of Natural Resources U.S. Geological Survey 125 Biological Sciences East National Park Service Tucson, Arizona 85721 U.S. Department of the Interior DIRK KEMPTHORNE, Secretary U.S. Geological Survey Mark Myers, Director U.S. Geological Survey, Reston, Virginia: 2008 For product and ordering information: World Wide Web: http://www.usgs.gov/pubprod Telephone: 1-888-ASK-USGS For more information on the USGS-the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment: World Wide Web:http://www.usgs.gov Telephone: 1-888-ASK-USGS Suggested Citation Powell, B.F., Schmidt, C.A., Halvorson, W.L., and Anning, Pamela, 2008, Vascular plant and vertebrate inventory of Chiricahua National Monument: U.S. Geological Survey Open-File Report 2008-1023, 104 p. [http://pubs.usgs.gov/of/2008/1023/]. Cover photo: Chiricahua National Monument. Photograph by National Park Service. Note: This report supersedes Schmidt et al. (2005). Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. -
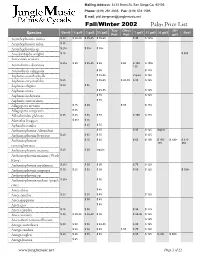
Winter-Fall Sale 2002 Palm Trees-Web
Mailing Address: 3233 Brant St. San Diego Ca, 92103 Phone: (619) 291 4605 Fax: (619) 574 1595 E mail: [email protected] Fall/Winter 2002 Palm Price List Tree Citrus 25/+ Band$ 1 gal$ 2 gal$ 3/5 gal$ 7 gal$ 15 gal$ 20 gal$ Box$ Species Pot$ Pot$ gal$ Acanthophoenix crinita $ 30 $ 30-40 $ 35-45 $ 55-65 $ 95 $ 125+ Acanthophoenix rubra $ 35 Acanthophoenix sp. $ 25+ $ 35+ $ 55+ Acoelorrhaphe wrightii $ 15 $ 300 Acrocomia aculeata $ 25+ $ 35 $ 35-45 $ 65 $ 65 $ 100- $ 150+ Actinokentia divaricata 135 Actinorhytis calapparia $ 55 $ 125 Aiphanes acanthophylla $ 45-55 inquire $ 125 Aiphanes caryotaefolia $ 25 $ 55-65 $ 45-55 $ 85 $ 125 Aiphanes elegans $ 20 $ 35 Aiphanes erosa $ 45-55 $ 125 Aiphanes lindeniana $ 55 $ 125 Aiphanes vincentsiana $ 55 Allagoptera arenaria $ 25 $ 40 $ 55 $ 135 Allagoptera campestris $ 35 Alloschmidtia glabrata $ 35 $ 45 $ 55 $ 85 $ 150 $ 175 Alsmithia longipes $ 35+ $ 55 Aphandra natalia $ 35 $ 55 Archontophoenix Alexandrae $ 55 $ 85 $ 125 inquire Archontophoenix Beatricae $ 20 $ 35 $ 55 $ 125 Archontophoenix $ 25 $ 45 $ 65 $ 100 $ 150- $ 200+ $ 310- 175 350 cunninghamiana Archontophoenix maxima $ 25 $ 30 inquire Archontophoenix maxima (Wash River) Archontophoenix myolaensis $ 25+ $ 30 $ 50 $ 75 $ 125 Archontophoenix purpurea $ 30 $ 25 $ 35 $ 50 $ 85 $ 125 $ 300+ Archontophoenix sp. Archontophoenix tuckerii (peach $ 25+ $ 55 river) Areca alicae $ 45 Areca catechu $ 20 $ 35 $ 45 $ 125 Areca guppyana $ 30 $ 45 Areca ipot $ 45 Areca triandra $ 25 $ 30 $ 95 $ 125 Areca vestiaria $ 25 $ 30-35 $ 35-40 $ 55 $ 85-95 $ 125 Arecastrum romanzoffianum $ 125 Arenga australasica $ 20 $ 30 $ 35 $ 45-55 $ 85 $ 125 Arenga caudata $ 20 $ 30 $ 45 $ 55 $ 75 $ 100 Arenga engleri $ 20 $ 60 $ 35 $ 45 $ 85 $ 125 $ 200 $ 300+ Arenga hastata $ 25 www.junglemusic.net Page 1 of 22 Tree Citrus 25/+ Band$ 1 gal$ 2 gal$ 3/5 gal$ 7 gal$ 15 gal$ 20 gal$ Box$ Species Pot$ Pot$ gal$ Arenga hookeriana inquire Arenga micranthe 'Lhutan' $ 20 inquire Arenga pinnata $ 35 $ 50 $ 85 $ 125 Arenga sp. -

Inventory of Taxa for the Fitzgerald River National Park
Flora Survey of the Coastal Catchments and Ranges of the Fitzgerald River National Park 2013 Damien Rathbone Department of Environment and Conservation, South Coast Region, 120 Albany Hwy, Albany, 6330. USE OF THIS REPORT Information used in this report may be copied or reproduced for study, research or educational purposed, subject to inclusion of acknowledgement of the source. DISCLAIMER The author has made every effort to ensure the accuracy of the information used. However, the author and participating bodies take no responsibiliy for how this informrion is used subsequently by other and accepts no liability for a third parties use or reliance upon this report. CITATION Rathbone, DA. (2013) Flora Survey of the Coastal Catchments and Ranges of the Fitzgerald River National Park. Unpublished report. Department of Environment and Conservation, Western Australia. ACKNOWLEDGEMENTS The author would like to thank many people that provided valable assistance and input into the project. Sarah Barrett, Anita Barnett, Karen Rusten, Deon Utber, Sarah Comer, Charlotte Mueller, Jason Peters, Roger Cunningham, Chris Rathbone, Carol Ebbett and Janet Newell provided assisstance with fieldwork. Carol Wilkins, Rachel Meissner, Juliet Wege, Barbara Rye, Mike Hislop, Cate Tauss, Rob Davis, Greg Keighery, Nathan McQuoid and Marco Rossetto assissted with plant identification. Coralie Hortin, Karin Baker and many other members of the Albany Wildflower society helped with vouchering of plant specimens. 2 Contents Abstract .............................................................................................................................. -

Primitive Angiosperm Flower – a Discussion*
Acta Bot. Neerl. 23(4), August 1974, p. 461-471. The structure and function of the primitive Angiosperm flower – a discussion* Gerhard Gottsberger Departamentode Botanica, Faculdade de Ciencias Medicas e Biologicas de Botucatu, Estado de Sao Paulo, Brazil SUMMARY Morphological and functional features of primitive entomophilous Angiosperm flowers are discussed and confronted with modem conceptions onearly Angiosperm differentiation. Evidence is put forward to show that large, solitary and terminally-borne flowers are not most primitive in the Angiosperms, but rather middle-sized ones, groupedinto lateral flower aggregates or inflorescences. It is believed that most primitive, still unspecialized Angiosperm flowers were pollinated casuallyby beetles. Only in a later phase did they graduallybecome adaptedto the more effective but more devastating type of beetle pollination. Together with this specialization, flower enlargment, reduction of inflorescences, numerical increase of stamens and carpels, and their more dense aggregationand flatteningmight have occurred. In have maintained the archaic condi- regard to pollination,many primitive Angiosperms tion of because beetles still dominant insect whereas in cantharophily, are a group, dispersal they have been largely forced to switch over from the archaic saurochory to the more modern modes of dispersal by birds and mammals,since duringthe later Mesozoic the dominance of reptiles had come to an end. The prevailing ideas regarding the primitiveness of Angiosperm flower struc- be somewhat Is the -

Plant Life of Western Australia
INTRODUCTION The characteristic features of the vegetation of Australia I. General Physiography At present the animals and plants of Australia are isolated from the rest of the world, except by way of the Torres Straits to New Guinea and southeast Asia. Even here adverse climatic conditions restrict or make it impossible for migration. Over a long period this isolation has meant that even what was common to the floras of the southern Asiatic Archipelago and Australia has become restricted to small areas. This resulted in an ever increasing divergence. As a consequence, Australia is a true island continent, with its own peculiar flora and fauna. As in southern Africa, Australia is largely an extensive plateau, although at a lower elevation. As in Africa too, the plateau increases gradually in height towards the east, culminating in a high ridge from which the land then drops steeply to a narrow coastal plain crossed by short rivers. On the west coast the plateau is only 00-00 m in height but there is usually an abrupt descent to the narrow coastal region. The plateau drops towards the center, and the major rivers flow into this depression. Fed from the high eastern margin of the plateau, these rivers run through low rainfall areas to the sea. While the tropical northern region is characterized by a wet summer and dry win- ter, the actual amount of rain is determined by additional factors. On the mountainous east coast the rainfall is high, while it diminishes with surprising rapidity towards the interior. Thus in New South Wales, the yearly rainfall at the edge of the plateau and the adjacent coast often reaches over 100 cm.