41 EUTHERIA METATHERIA Tree of Mammals
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Dated Phylogeny of Marsupials Using a Molecular Supermatrix and Multiple Fossil Constraints
Journal of Mammalogy, 89(1):175–189, 2008 A DATED PHYLOGENY OF MARSUPIALS USING A MOLECULAR SUPERMATRIX AND MULTIPLE FOSSIL CONSTRAINTS ROBIN M. D. BECK* School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, New South Wales 2052, Australia Downloaded from https://academic.oup.com/jmammal/article/89/1/175/1020874 by guest on 25 September 2021 Phylogenetic relationships within marsupials were investigated based on a 20.1-kilobase molecular supermatrix comprising 7 nuclear and 15 mitochondrial genes analyzed using both maximum likelihood and Bayesian approaches and 3 different partitioning strategies. The study revealed that base composition bias in the 3rd codon positions of mitochondrial genes misled even the partitioned maximum-likelihood analyses, whereas Bayesian analyses were less affected. After correcting for base composition bias, monophyly of the currently recognized marsupial orders, of Australidelphia, and of a clade comprising Dasyuromorphia, Notoryctes,and Peramelemorphia, were supported strongly by both Bayesian posterior probabilities and maximum-likelihood bootstrap values. Monophyly of the Australasian marsupials, of Notoryctes þ Dasyuromorphia, and of Caenolestes þ Australidelphia were less well supported. Within Diprotodontia, Burramyidae þ Phalangeridae received relatively strong support. Divergence dates calculated using a Bayesian relaxed molecular clock and multiple age constraints suggested at least 3 independent dispersals of marsupials from North to South America during the Late Cretaceous or early Paleocene. Within the Australasian clade, the macropodine radiation, the divergence of phascogaline and dasyurine dasyurids, and the divergence of perameline and peroryctine peramelemorphians all coincided with periods of significant environmental change during the Miocene. An analysis of ‘‘unrepresented basal branch lengths’’ suggests that the fossil record is particularly poor for didelphids and most groups within the Australasian radiation. -

A Phylogeny and Timescale for Marsupial Evolution Based on Sequences for Five Nuclear Genes
J Mammal Evol DOI 10.1007/s10914-007-9062-6 ORIGINAL PAPER A Phylogeny and Timescale for Marsupial Evolution Based on Sequences for Five Nuclear Genes Robert W. Meredith & Michael Westerman & Judd A. Case & Mark S. Springer # Springer Science + Business Media, LLC 2007 Abstract Even though marsupials are taxonomically less diverse than placentals, they exhibit comparable morphological and ecological diversity. However, much of their fossil record is thought to be missing, particularly for the Australasian groups. The more than 330 living species of marsupials are grouped into three American (Didelphimorphia, Microbiotheria, and Paucituberculata) and four Australasian (Dasyuromorphia, Diprotodontia, Notoryctemorphia, and Peramelemorphia) orders. Interordinal relationships have been investigated using a wide range of methods that have often yielded contradictory results. Much of the controversy has focused on the placement of Dromiciops gliroides (Microbiotheria). Studies either support a sister-taxon relationship to a monophyletic Australasian clade or a nested position within the Australasian radiation. Familial relationships within the Diprotodontia have also proved difficult to resolve. Here, we examine higher-level marsupial relationships using a nuclear multigene molecular data set representing all living orders. Protein-coding portions of ApoB, BRCA1, IRBP, Rag1, and vWF were analyzed using maximum parsimony, maximum likelihood, and Bayesian methods. Two different Bayesian relaxed molecular clock methods were employed to construct a timescale for marsupial evolution and estimate the unrepresented basal branch length (UBBL). Maximum likelihood and Bayesian results suggest that the root of the marsupial tree is between Didelphimorphia and all other marsupials. All methods provide strong support for the monophyly of Australidelphia. Within Australidelphia, Dromiciops is the sister-taxon to a monophyletic Australasian clade. -

Amblysomus Robustus – Robust Golden Mole
Amblysomus robustus – Robust Golden Mole continuing decline and possible severe fragmentation of habitat. Currently known from only five locations but probably more widespread. Further field surveys and molecular data are needed to accurately delimit its range. The Highveld grasslands favoured by this species are being degraded by mining for shallow coal deposits to fuel numerous power stations that occur in the preferred high-altitude grassland habitats of this species, which is an inferred major threat. Rehabilitation attempts at these sites appear to have been largely ineffective. These power stations form the backbone of South Africa's electricity network, and disturbance is likely to increase as human populations grow and the demand for power increases. While no mining sites and power generation plants occur at the five localities where this species has been collected, an environmental authorisation application to mine coal at a site near Belfast, close to where this species occurs, is Gary Bronner currently being assessed. Given the ubiquity of mines and power stations in the Mpumalanga grasslands, impacts on this species are likely if it is more widespread than current Regional Red List status (2016) Vulnerable B1ab(iii)* records indicate, which seems probable. Farming, tourism National Red List status (2004) Endangered resort developments and agro-forestry (exotic pine and B1,2ab(i-iv) eucalyptus plantations) have also transformed habitat, but less dramatically; these do not appear to pose a major Reasons for change Non-genuine: threat. More data is required on the distribution limits, New information ecology, densities and reproduction of this species. Global Red List status (2015) Vulnerable B1ab(iii) TOPS listing (NEMBA) None Distribution CITES listing None Endemic to South Africa, this species is known from only the Steenkampsberg Mountain Plateau and in the Endemic Yes Dullstroom and Belfast areas of Mpumalanga (Figure 1), extending eastwards to Lydenburg and possibly *Watch-list Data southwards towards the Ermelo district where A. -

Perissodactyla: Tapirus) Hints at Subtle Variations in Locomotor Ecology
JOURNAL OF MORPHOLOGY 277:1469–1485 (2016) A Three-Dimensional Morphometric Analysis of Upper Forelimb Morphology in the Enigmatic Tapir (Perissodactyla: Tapirus) Hints at Subtle Variations in Locomotor Ecology Jamie A. MacLaren1* and Sandra Nauwelaerts1,2 1Department of Biology, Universiteit Antwerpen, Building D, Campus Drie Eiken, Universiteitsplein, Wilrijk, Antwerp 2610, Belgium 2Centre for Research and Conservation, Koninklijke Maatschappij Voor Dierkunde (KMDA), Koningin Astridplein 26, Antwerp 2018, Belgium ABSTRACT Forelimb morphology is an indicator for order Perissodactyla (odd-toed ungulates). Modern terrestrial locomotor ecology. The limb morphology of the tapirs are widely accepted to belong to a single enigmatic tapir (Perissodactyla: Tapirus) has often been genus (Tapirus), containing four extant species compared to that of basal perissodactyls, despite the lack (Hulbert, 1973; Ruiz-Garcıa et al., 1985) and sev- of quantitative studies comparing forelimb variation in eral regional subspecies (Padilla and Dowler, 1965; modern tapirs. Here, we present a quantitative assess- ment of tapir upper forelimb osteology using three- Wilson and Reeder, 2005): the Baird’s tapir (T. dimensional geometric morphometrics to test whether bairdii), lowland tapir (T. terrestris), mountain the four modern tapir species are monomorphic in their tapir (T. pinchaque), and the Malayan tapir (T. forelimb skeleton. The shape of the upper forelimb bones indicus). Extant tapirs primarily inhabit tropical across four species (T. indicus; T. bairdii; T. terrestris; T. rainforest, with some populations also occupying pinchaque) was investigated. Bones were laser scanned wet grassland and chaparral biomes (Padilla and to capture surface morphology and 3D landmark analysis Dowler, 1965; Padilla et al., 1996). was used to quantify shape. -

Controlled Animals
Environment and Sustainable Resource Development Fish and Wildlife Policy Division Controlled Animals Wildlife Regulation, Schedule 5, Part 1-4: Controlled Animals Subject to the Wildlife Act, a person must not be in possession of a wildlife or controlled animal unless authorized by a permit to do so, the animal was lawfully acquired, was lawfully exported from a jurisdiction outside of Alberta and was lawfully imported into Alberta. NOTES: 1 Animals listed in this Schedule, as a general rule, are described in the left hand column by reference to common or descriptive names and in the right hand column by reference to scientific names. But, in the event of any conflict as to the kind of animals that are listed, a scientific name in the right hand column prevails over the corresponding common or descriptive name in the left hand column. 2 Also included in this Schedule is any animal that is the hybrid offspring resulting from the crossing, whether before or after the commencement of this Schedule, of 2 animals at least one of which is or was an animal of a kind that is a controlled animal by virtue of this Schedule. 3 This Schedule excludes all wildlife animals, and therefore if a wildlife animal would, but for this Note, be included in this Schedule, it is hereby excluded from being a controlled animal. Part 1 Mammals (Class Mammalia) 1. AMERICAN OPOSSUMS (Family Didelphidae) Virginia Opossum Didelphis virginiana 2. SHREWS (Family Soricidae) Long-tailed Shrews Genus Sorex Arboreal Brown-toothed Shrew Episoriculus macrurus North American Least Shrew Cryptotis parva Old World Water Shrews Genus Neomys Ussuri White-toothed Shrew Crocidura lasiura Greater White-toothed Shrew Crocidura russula Siberian Shrew Crocidura sibirica Piebald Shrew Diplomesodon pulchellum 3. -
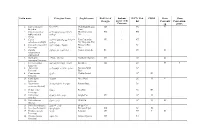
Red List of Endemic IUCN Red CITES Bern Bonn Georgia Species of the List Conventi Convention Caucasus on (CMS) 1
Latin name Georgian Name English name Red List of Endemic IUCN Red CITES Bern Bonn Georgia species of the list Conventi Convention Caucasus on (CMS) 1. Capra aegagrus niamori Wild Goat, Bezoar CR VU II Erxleben. Goat 2. Capra caucasica dasavleTkavkasiuri West Caucasian EN + EN Güldenstädt & jixvi Tur Pallas. 3. Capra aRmosavleTkavkasiuri East Caucasian VU + NT cylindricornis Blyth. jixvi Tur, Dagestan Tur 4. Capreolus capreolus evropuli Sveli European Roe LC Linnaeus. Deer 5. Gazella qurciki, jeirani Goitered Gazelle RE VU II subgutturosa Güldenstädt. 6. Rupicapra arCvi, fsiti Northern Chamois EN LC II rupicapra Linnaeus. 7. Cervus elaphus keTilSobili iremi Red Deer CR LC II I Linnaeus. 8. Sus scrofa gareuli Rori, taxi Eurasian Wild LC Linnaeus. Boar 9. Canis aureus tura Golden Jackal LC III Linnaeus. 10. Canis lupus mgeli Grey Wolf LC II II Linnaeus. 11. Nyctereutes enotisebri ZaRli Racoon Dog LC procyonoides Gray. 12. Vulpes vulpes mela Red Fox LC III Linnaeus. 13. Felis chaus lelianis kata Jungle Cat VU LC II Schreber. 14. Felis silvestris tyis kata Wild Cat LC II II Shreber. 15. Felis libyca Forster. velis kata Steppe Cat 16. Lynx lynx Linnaeus. focxveri Eurasian Lynx CR LC II 17. Panthera pardus jiqi Leopard CR NT I II Linnaeus. 18. Hyaena hyaena afTari Striped Hyaena CR NT Linnaeus. 19. Lutra lutra wavi Eurasian Otter, VU NT I II Linnaeus. Common Otter 20. Martes foina kldis kverna Stone Marten, LC III Erxleben. Beech Marten 21. Martes martes tyis kverna European Pine LC Linnaeus. Marten 22. Meles meles maCvi Eurasian Badger LC Linnaeus. 23. Mustela lutreola waula European Mink EN II Linnaeus. -

Miocene Mammal Reveals a Mesozoic Ghost Lineage on Insular New Zealand, Southwest Pacific
Miocene mammal reveals a Mesozoic ghost lineage on insular New Zealand, southwest Pacific Trevor H. Worthy*†, Alan J. D. Tennyson‡, Michael Archer§, Anne M. Musser¶, Suzanne J. Hand§, Craig Jonesʈ, Barry J. Douglas**, James A. McNamara††, and Robin M. D. Beck§ *School of Earth and Environmental Sciences, Darling Building DP 418, Adelaide University, North Terrace, Adelaide 5005, South Australia, Australia; ‡Museum of New Zealand Te Papa Tongarewa, P.O. Box 467, Wellington 6015, New Zealand; §School of Biological, Earth and Environmental Sciences, University of New South Wales, New South Wales 2052, Australia; ¶Australian Museum, 6-8 College Street, Sydney, New South Wales 2010, Australia; ʈInstitute of Geological and Nuclear Sciences, P.O. Box 30368, Lower Hutt 5040, New Zealand; **Douglas Geological Consultants, 14 Jubilee Street, Dunedin 9011, New Zealand; and ††South Australian Museum, Adelaide, South Australia 5000, Australia Edited by James P. Kennett, University of California, Santa Barbara, CA, and approved October 11, 2006 (sent for review July 8, 2006) New Zealand (NZ) has long been upheld as the archetypical Ma) dinosaur material (13) and isolated moa bones from marine example of a land where the biota evolved without nonvolant sediments up to 2.5 Ma (1, 14), the terrestrial record older than terrestrial mammals. Their absence before human arrival is mys- 1 Ma is extremely limited. Until now, there has been no direct terious, because NZ was still attached to East Antarctica in the Early evidence for the pre-Pleistocene presence in NZ of any of its Cretaceous when a variety of terrestrial mammals occupied the endemic vertebrate lineages, particularly any group of terrestrial adjacent Australian portion of Gondwana. -

A Species-Level Phylogenetic Supertree of Marsupials
J. Zool., Lond. (2004) 264, 11–31 C 2004 The Zoological Society of London Printed in the United Kingdom DOI:10.1017/S0952836904005539 A species-level phylogenetic supertree of marsupials Marcel Cardillo1,2*, Olaf R. P. Bininda-Emonds3, Elizabeth Boakes1,2 and Andy Purvis1 1 Department of Biological Sciences, Imperial College London, Silwood Park, Ascot SL5 7PY, U.K. 2 Institute of Zoology, Zoological Society of London, Regent’s Park, London NW1 4RY, U.K. 3 Lehrstuhl fur¨ Tierzucht, Technical University of Munich, Alte Akademie 12, 85354 Freising-Weihenstephan, Germany (Accepted 26 January 2004) Abstract Comparative studies require information on phylogenetic relationships, but complete species-level phylogenetic trees of large clades are difficult to produce. One solution is to combine algorithmically many small trees into a single, larger supertree. Here we present a virtually complete, species-level phylogeny of the marsupials (Mammalia: Metatheria), built by combining 158 phylogenetic estimates published since 1980, using matrix representation with parsimony. The supertree is well resolved overall (73.7%), although resolution varies across the tree, indicating variation both in the amount of phylogenetic information available for different taxa, and the degree of conflict among phylogenetic estimates. In particular, the supertree shows poor resolution within the American marsupial taxa, reflecting a relative lack of systematic effort compared to the Australasian taxa. There are also important differences in supertrees based on source phylogenies published before 1995 and those published more recently. The supertree can be viewed as a meta-analysis of marsupial phylogenetic studies, and should be useful as a framework for phylogenetically explicit comparative studies of marsupial evolution and ecology. -

Convergent Evolution in the Euarchontoglires
This is a repository copy of Convergent evolution in the Euarchontoglires. White Rose Research Online URL for this paper: https://eprints.whiterose.ac.uk/133262/ Version: Published Version Article: Morris, Philip James Rencher, Cobb, Samuel Nicholas Frederick orcid.org/0000-0002- 8360-8024 and Cox, Philip Graham orcid.org/0000-0001-9782-2358 (2018) Convergent evolution in the Euarchontoglires. Biology letters. 2018036. ISSN 1744-957X https://doi.org/10.1098/rsbl.2018.0366 Reuse This article is distributed under the terms of the Creative Commons Attribution (CC BY) licence. This licence allows you to distribute, remix, tweak, and build upon the work, even commercially, as long as you credit the authors for the original work. More information and the full terms of the licence here: https://creativecommons.org/licenses/ Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ Downloaded from http://rsbl.royalsocietypublishing.org/ on August 1, 2018 Evolutionary biology Convergent evolution in the rsbl.royalsocietypublishing.org Euarchontoglires Philip J. R. Morris1, Samuel N. F. Cobb2 and Philip G. Cox2 1Hull York Medical School, University of Hull, Hull HU6 7RX, UK Research 2Department of Archaeology and Hull York Medical School, University of York, York YO10 5DD, UK SNFC, 0000-0002-8360-8024; PGC, 0000-0001-9782-2358 Cite this article: Morris PJR, Cobb SNF, Cox PG. 2018 Convergent evolution in the Convergence—the independent evolution of similar phenotypes in distantly Euarchontoglires. -

The Impact of Locomotion on the Brain Evolution of Squirrels and Close Relatives ✉ Ornella C
ARTICLE https://doi.org/10.1038/s42003-021-01887-8 OPEN The impact of locomotion on the brain evolution of squirrels and close relatives ✉ Ornella C. Bertrand 1 , Hans P. Püschel 1, Julia A. Schwab 1, Mary T. Silcox 2 & Stephen L. Brusatte1 How do brain size and proportions relate to ecology and evolutionary history? Here, we use virtual endocasts from 38 extinct and extant rodent species spanning 50+ million years of evolution to assess the impact of locomotion, body mass, and phylogeny on the size of the brain, olfactory bulbs, petrosal lobules, and neocortex. We find that body mass and phylogeny are highly correlated with relative brain and brain component size, and that locomotion strongly influences brain, petrosal lobule, and neocortical sizes. Notably, species living in 1234567890():,; trees have greater relative overall brain, petrosal lobule, and neocortical sizes compared to other locomotor categories, especially fossorial taxa. Across millions of years of Eocene- Recent environmental change, arboreality played a major role in the early evolution of squirrels and closely related aplodontiids, promoting the expansion of the neocortex and petrosal lobules. Fossoriality in aplodontiids had an opposing effect by reducing the need for large brains. 1 School of GeoSciences, University of Edinburgh, Grant Institute, Edinburgh, Scotland, UK. 2 Department of Anthropology, University of Toronto Scarborough, ✉ Toronto, ON, Canada. email: [email protected] COMMUNICATIONS BIOLOGY | (2021) 4:460 | https://doi.org/10.1038/s42003-021-01887-8 | www.nature.com/commsbio 1 ARTICLE COMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01887-8 hat ecological and evolutionary factors affect brain size striking differences between living sciurids and aplodontiids raise in mammals? Studies have assessed the impact of the question of how this modern rodent assemblage emerged. -

Relational Database Systems 1
Relational Database Systems 1 Wolf-Tilo Balke Jan-Christoph Kalo Institut für Informationssysteme Technische Universität Braunschweig www.ifis.cs.tu-bs.de Summary last week • Data models define the structural constrains and possible manipulations of data – Examples of Data Models: • Relational Model, Network Model, Object Model, etc. – Instances of data models are called schemas • Careful: Often, sloppy language is used where people call a schema also a model • We have three types of schemas: – Conceptual Schemas – Logical Schemas – Physical Schemas • We can use ER modeling for conceptual and logical schemas Relational Database Systems 1 – Wolf-Tilo Balke – Institut für Informationssysteme – TU Braunschweig 2 Summary last week • Entity Type Name • Weak Entity Type Name • Attribute name • Key Attribute name • name Multi-valued Attribute name name • Composite Attribute name • Derived Attribute name • Relationship Type name • Identifying Relationship Type name EN 3.5 Relational Database Systems 1 – Wolf-Tilo Balke – Institut für Informationssysteme – TU Braunschweig 3 Summary last week • Total participation of E2 in R E1 r E2 • Cardinality – an instance of E1 may relate to multiple instances of E2 (0,*) (1,1) E1 r E2 • Specific cardinality with min and max – an instance of E1 may relate to multiple instances of E2 (0,*) (0,1) E1 r E2 EN 3.5 Relational Database Systems 1 – Wolf-Tilo Balke – Institut für Informationssysteme – TU Braunschweig 4 3 Extended Data Modeling • Alternative ER Notations • Extended ER – Inheritance – Complex Relationships -

The Adapted Ears of Big Cats and Golden Moles: Exotic Outcomes of the Evolutionary Radiation of Mammals
FEATURED ARTICLE The Adapted Ears of Big Cats and Golden Moles: Exotic Outcomes of the Evolutionary Radiation of Mammals Edward J. Walsh and JoAnn McGee Through the process of natural selection, diverse organs and organ systems abound throughout the animal kingdom. In light of such abundant and assorted diversity, evolutionary adaptations have spawned a host of peculiar physiologies. The anatomical oddities that underlie these physiologies and behaviors are the telltale indicators of trait specialization. Following from this, the purpose of this article is to consider a number of auditory “inventions” brought about through natural selection in two phylogenetically distinct groups of mammals, the largely fossorial golden moles (Order Afrosoricida, Family Chrysochloridae) and the carnivorous felids of the genus Panthera along with its taxonomic neigh- bor, the clouded leopard (Neofelis nebulosa). In the Beginning The first vertebrate land invasion occurred during the Early Carboniferous period some 370 million years ago. The primitive but essential scaffolding of what would become the middle and inner ears of mammals was present at this time, although the evolution of the osseous (bony) middle ear system and the optimization of cochlear fea- tures and function would play out over the following 100 million years. Through natural selection, the evolution of the middle ear system, composed of three small articu- lated bones, the malleus, incus, and stapes, and a highly structured and coiled inner ear, came to represent all marsupial and placental (therian) mammals on the planet Figure 1. Schematics of the outer, middle, and inner ears (A) and thus far studied. The consequences of this evolution were the organ of Corti in cross section (B) of a placental mammal.