Utrecht Micropaleontological Bulletins
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Chania : Explore & Experience
INDEX INDEX .......................................................................................................................................... 1 THE BYZANTINE WALL OF CHANIA ............................................................................................. 3 THE EGYPTIAN LIGHTHOUSE ...................................................................................................... 4 GIALI TZAMISI ............................................................................................................................. 5 VENETIAN NEORIA ...................................................................................................................... 6 FIRKA FORTRESS ......................................................................................................................... 7 CENTER OF MEDITERRANEAN ARCHITECTURE (GRAND ARSENAL)............................................ 8 ANCIENT KYDONIA (PROTO-MINOAN SETTLEMENT OF KASTELI) .............................................. 9 ANCIENT APTERA ......................................................................................................................10 ENTRANCE OF THE RENIER MANSION ......................................................................................11 GATE AND RAMPART SABBIONARA .........................................................................................12 THE MINARET OF AGIOS NIKOLAOS .........................................................................................13 THE GRAVES OF VENIZELOS FAMILY ........................................................................................14 -

Case Study #5: the Myrtoon Sea/ Peloponnese - Crete
Addressing MSP Implementation in Case Study Areas Case Study #5: The Myrtoon Sea/ Peloponnese - Crete Passage Deliverable C.1.3.8. Co-funded by the1 European Maritime and Fisheries Fund of the European Union. Agreement EASME/EMFF/2015/1.2.1.3/01/S12.742087 - SUPREME ACKNOWLEDGEMENT The work described in this report was supported by the European Maritime and Fisheries Fund of the European Union- through the Grant Agreement EASME/EMFF/2015/1.2.1.3/01/S12.742087 - SUPREME, corresponding to the Call for proposal EASME/EMFF/2015/1.2.1.3 for Projects on Maritime Spatial Planning (MSP). DISCLAIMERS This document reflects only the authors’ views and not those of the European Union. This work may rely on data from sources external to the SUPREME project Consortium. Members of the Consortium do not accept liability for loss or damage suffered by any third party as a result of errors or inaccuracies in such data. The user thereof uses the information at its sole risk and neither the European Union nor any member of the SUPREME Consortium, are liable for any use that may be made of the information The designations employed and the presentation of material in the present document do not imply the expression of any opinion on the part of UN Environment/MAP Barcelona Convention Secretariat concerning the legal status of any country, territory, area, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The depiction and use of boundaries, geographic names and related data shown on maps included in the present document are not warranted to be error free nor do they imply official endorsement or acceptance by UN Environment/ MAP Barcelona Convention Secretariat. -

Alternative Spatial Arrangement of Small-Sized Municipal Solid Waste Bins
ALTERNATIVE SPATIAL ARRANGEMENT OF SMALL-SIZED MUNICIPAL SOLID WASTE BINS I. KARAGIANNAKI, E. GIDARAKOS SCHOOL OF ENVIRONMENTAL ENGINEERING, TECHNICAL UNIVERSITY OF CRETE, POLITECHNIOUPOLIS 73100 CHANIA, GREECE SUMMARY: Solid waste management stategy that every cosiety adopts is of the utmost meaning. The object of this study was to evaluate quantitative and qualitative the existing solid waste management of survey area, which is Old Harbor of Chania, Crete. In terms of quantity, the max total capacity of small-sized solid waste bins adequates to cover the demands of study area's visitors. Unfortunately, in terms of quality, it has been observed many points devastate the ergonomy and safety of urban surroundings. So, without changes in operating cost, two different scenarios of alternative spacial arrangement of small-sized municipal solid waste bins were designed, compling with principles of Neüfert (Ernst Neüfert, 2003). According to results, both of scenarios improve the servicing level of area, in comparison with the existing situation. However, in first scenario it ιs observed bigger dispersion of bins not only in central roads but in these with medium traffic too. 1. INTRODUCTION Increasing evolution of society affect and definately will continue to influence the prodused solid wastes not only quantitatively but also qualitatively (Liolios, G.2014). However, countries has adopted different solid wastes management strategies to maintain the high standarts of public hygiene. The basic stages of a solid waste management system are waste generation, waste disposal, collection, transport, treatment, recovery and disposal in landfills (Rada, E. Et al.2013). The abundance of these systems is the result of different economic, climatic, geomorpfological and cultural parameters that characterize each society επίπεδο (Tseridou, Ε. -

Thehotel.Gr Travel Agency, +30 2821090760, Chania – Crete, Guide of Crete
TheHotel.gr Travel Agency, +30 2821090760, Chania – Crete, Guide of Crete TheHotel.gr Travel Agency Chania – Crete – Greece 41-43 Skalidi street, Chania +30 2821090760 [email protected] Guide of Crete http://www.thehotel.gr 0 TheHotel.gr Travel Agency, +30 2821090760, Chania – Crete, Guide of Crete CONTENTS CONTENTS ....................................................................................... 1 1. ABOUT THEHOTEL.GR TRAVEL AGENCY ........................................ 3 2. WHY BOOK WITH US .................................................................... 4 OUR COMPANY ........................................................................................ 4 YOU ARE OUR PRIORITY ......................................................................... 4 PERSONALIZED SERVICE ......................................................................... 4 EMPOWERMENT ....................................................................................... 4 SAVINGS ................................................................................................. 4 BOOK WITH CONFIDENCE AND SECURITY ............................................... 5 3. GREECE ........................................................................................ 6 4. CRETE .......................................................................................... 7 AREAS OF NATURAL BEAUTY ................................................................. 10 1. The White Mountains, Chania area ....................................................... 10 2. -

Chania Premium Sunset Safari Tour
Chania Premium Sunset Safari Tour... Highlights Pick Up Times Schedule 17:15 Chania May - October: Monday - Thursday 17:15 Nea Chora 17:25 Golden Beach Price Duration Distance 17:25 Agioi Apostoloi 72.00€Adults and Children 4 - 5 Hours 70 Km 17:30 Kalamaki The prices are for group participation with minimum 17:35 Stalos of 4 pax to start. 17:40 Agia Marina 17:45 Platanias 17:50 Gerani Price Includes Difficulty Altitude Dif: 17:55 Maleme Free hotel pick up and drop off. 3.0 700m 18:00 Tavronitis Premium offroad vehicles (7 or 9 seater) driven by 18:05 Kolimvary multilingual escort-drivers Semi private tour up to 6 or 8 persons per vehicle Three Wines tasting with Cretan delicacies platter. Sunset panoramic view of Chania Liability insurance and local taxes. Uncharted off road routes in Non-touristic areas Traditional villages Wine Tasting Gallery 1 Price Does Not Include Extra wine consumption Gratuities Description Premium Sunset Safari Tour in the Northwest side of the inland of Crete... Allow yourself to indulge in a romantic off-road route experience with the best panoramic view followed by a wine tasting with delicacies platter in a semi private tour with up to 6 or 8 persons per vehicle. Our evening tour begins from Chania city centre and tourist areas, heading towards the White Mountains through uncharted mountain routes. Our premium off-road vehicles driven by well trained escorts will help you enjoy the best panoramic sunset view of Northwest Crete from above. Unpaved off-road routes, with wild and dense vegetation, non tourist places, unique panoramic views, ideal for sunset capture stops, combined with a three wines tasting accompanied by Cretan delicacies platter, are only some of the experiences that you will have in this 4-5 hour premium sunset safari tour. -

On the Electric Field Transient Anomaly Observed at the Time of the Kythira
Nat. Hazards Earth Syst. Sci., 7, 677–682, 2007 www.nat-hazards-earth-syst-sci.net/7/677/2007/ Natural Hazards © Author(s) 2007. This work is licensed and Earth under a Creative Commons License. System Sciences On the electric field transient anomaly observed at the time of the Kythira M=6.9 earthquake on January 2006 A. Konstantaras1, J. P. Makris1, F. Vallianatos1, and M. R. Varley2 1Technological Educational Institute of Crete, Chania, Crete, 731 33, Greece 2ADSIP Research Centre, Department of Technology, University of Central Lancashire, Preston, PR1 2HE, UK Received: 10 July 2007 – Revised: 29 October 2007 – Accepted: 29 October 2007 – Published: 22 November 2007 Abstract. The study of the Earth’s electromagnetic fields the INTAS-99-1102 project titled “Study of the ULF elec- prior to the occurrence of strong seismic events has repeat- tromagnetic phenomena related to earthquakes (SUPRE)”. edly revealed cases were transient anomalies, often deemed The MVC-2DS recording station (Hattori et al., 2004) has as possible earthquake precursors, were observed on electro- the capacity to measure three electric (telluric) components magnetic field recordings of surface, atmosphere and near (dynamic range ±2.5 V) and uses a torsion photoelectric space carried out measurements. In an attempt to under- magnetometer (frequency range ∼15 Hz, measurement range stand the nature of such signals several models have been ±5000 nT). The sampling frequency is 50 Hz. The magnetic proposed based upon the exhibited characteristics of the ob- sensor is installed in the ground at 0.5 m depth and is covered served anomalies and different possible generation mecha- with a plastic waterproof box. -

Landslide Mapping and Susceptibility Assessment Using Geospatial Analysis and Earth Observation Data
land Article Landslide Mapping and Susceptibility Assessment Using Geospatial Analysis and Earth Observation Data Emmanouil Psomiadis 1,* , Andreas Papazachariou 1, Konstantinos X. Soulis 1 , Despoina-Simoni Alexiou 1 and Ioannis Charalampopoulos 2 1 Department of Natural Resources Management and Agricultural Engineering, Agricultural University of Athens, 75 Iera Odos st., 11855 Athens, Greece; [email protected] (A.P.); [email protected] (K.X.S.); [email protected] (D.-S.A.) 2 Department of Crop Science, Agricultural University of Athens, 75 Iera Odos st., 11855 Athens, Greece; [email protected] * Correspondence: [email protected]; Tel.: +30-210-529-4156 Received: 27 March 2020; Accepted: 27 April 2020; Published: 28 April 2020 Abstract: The western part of Crete Island has undergone serious landslide events in the past. The intense rainfalls that took place in the September 2018 to February 2019 period provoked extensive landslide events at the northern part of Chania prefecture, along the motorway A90. Geospatial analysis methods and earth observation data were utilized to investigate the impact of the various physical and anthropogenic factors on landslides and to evaluate landslide susceptibility. The landslide inventory map was created based on literature, aerial photo analysis, satellite images, and field surveys. A very high-resolution Digital Elevation Model (DEM) and land cover map was produced from a dense point cloud and Earth Observation data (Landsat 8), accordingly. Sentinel-2 data were used for the detection of the recent landslide events and offered suitable information for two of them. Eight triggering factors were selected and manipulated in a GIS-based environment. A semi-quantitative method of Analytical Hierarchy Process (AHP) and Weighted Linear Combination (WLC) was applied to evaluate the landslide susceptibility index (LSI) both for Chania prefecture and the motorway A90 in Chania. -

ARCHELON 2019 Crete Short Report Final
ARCHELON, the Sea Turtle Protection Society of Greece Conservation efforts during 2019 at the nesting habitats of Caretta caretta in Crete, Greece (Natura 2000 sites – GR4330004, GR4340003, GR4340006, GR4310004 and GR4310012) Athens, December 2019 1 Copyright © 2019 by ARCHELON No part of this report may be reproduced in any form or by any means without the written permission of ARCHELON ***************************************************************** ARCHELON, the Sea Turtle Protection Society of Greece Solomou 57 GR-104 32 ATHENS Greece Tel./Fax: +30-210-5231342 e-mail: [email protected] URL: www.archelon.gr 2 Contents Prologue p. 4 1. Background p. 4 1.1. Location and protection status of the nesting habitats p. 4 1.2. The long-term field projects of ARCHELON p. 4 2. The actual situation in the field during the 2019 nesting season p. 5 2.1. Brief description of the nesting sites p. 5 2.2. Anthropogenic disturbances on nesting beaches and the surrounding marine areas during 2019 p. 6 3. Conclusions and Recommendations p. 8 Illustrations p. 10 Photos p. 12 3 Prologue The aim of this short report is to present the major events in the efforts to protect the Loggerhead sea turtle’s ( Caretta caretta ) nesting habitats in Crete, as recorded through the fieldwork of ARCHELON. In particular, it includes the conservation status of the nesting areas at Rethymno Bay, Chania Bay and Messaras Bay, which are included in the Natura 2000 network under the site codes GR4330004 “PRASSANO FARANGI - PATSOS - SFAKORYAKO REMA - PARALIA RETHYMNOU KAI EKVOLI GEROPOTAMOU, AKR. LIANOS KAVOS – PERIVOLIA”, GR4340003 “CHERSONISOS RODOPOU - PARALIA MALEME”, GR4340006 “ LIMNI AGIAS - PLATANIAS - REMA KAI EKVOLI KERITI - KOILADA FASA ”, GR4310004 “DYTIKA ASTEROUSIA (APO AGIOFARANGO EOS KOKKINO PYRGO)” and GR4310012 “EKVOLI GEROPOTAMOU MESARAS”. -

In Tro D Uction I
1 i Introduction Incredible beachesIncredible Intro If someone asked you to list the reasons why you would choose a place to spend your summer vacation, what would those be? Don’t think about it too much. The trick is to give spontaneous an- swers, or even better, to mentally travel to that destination, create an image, place yourself in it at a central spot and describe it out loud. Let me guess: you already feel the sun warm on your face! You listen to the sound of the sea as if you held a shell to your ear. You have a camera in one hand, a cool cocktail in the other and you taste unprecedented gastronomical joys. Don’t open your eyes yet! Let a warm smile see you off just before you leave your imaginative tour and listen closely to that little whisper inside your mind… Discover the magic of Chania! 1 4 Reason No1 Incredible beachesIncredible Beach index It might sound like a truism when a tour- ist guide advertising a summer destina- tion refers to its beaches as “incredible”, but when it comes to Chania this is in fact the truth. Whether you are looking for, busy beaches with all kind of amenities or isolated paradises on earth, it is there! All you have to do is look for it… Elafonissi 8 Balos 14 Falassarna 20 Platanias 24 Stalos 54 Agia Marina 55 Chrissi Akti 62 Kalamaki 66 Platanias 68 Kolympari 72 Kasteli 74 Stavros 76 Kalyves 82 Georgioupolis 84 South Beaches 86 Palaiochora 88 2 8 Reason No1 Reason No1 9 Elafonissi Incredible beachesIncredible beachesIncredible If someone asked you til the island and feel like a which is your happiest castaway and a conqueror childhood memory, you at the same time! would probably include, “a Besides, the south coast day at the beach splashing of the island has many hid- my feet in the shallow wa- den beauties and pleas- ters of the sea”. -

15.80 EUR/Passenger/Way 16.90 EUR/Passenger/Way 14.70 EUR
Service is available for flights between 1st April and 31 st October CRETE CHANIA AIRPORT – CHANIA town and surroundings Shared door to door Service HOTELS ONLY! Chania town 14.70 EUR/passenger/way Daratso / Kato Galtatas / Ag. Apostoli/ Chrissi Akti / 15.80 EUR/passenger/way Kalamaki* (Chania) Platanias / Agia Marina / Stalos / Stavros / Akrotiri 16.90 EUR/passenger/way *no service to Kalamaki (Tympaki) Please note some hotels are not accessible by bus, in these cases you will be dropped of/picked up at a central point: Platanias area: Platanias square, EKO Gas Station Agia Marina area: Bus stop across Alexandra Beach hotel Kato Stalos area: CTS Office Agioi Apostoloi area: Traffic light across SYN.KA Super Market Chania Town: KTEL bus station (Central public bus station), Agora (Municipal Market of Chania), Alpha Bank at Halidon str. (Pick up point for all the hotels in the old harbor that are not accessible by bus) Meeting points: From Crete Chania Airport: At the airport our meeting point is in the Arrivals Hall. From city: In the city our driver will wait you at the hotel reception. Shared door-to-door transfer to Crete destinations: Our city shuttle service is a shared service – we pick-up/drop-off passengers at different locations. Journey time: approximately 30-100 minutes, depending on the traffic. It is important that the booked transfer gets only confirmed if the correct destination/pick-up address is given (we accept addresses only within the served area). Bookings with not full or with no destination/pick up address will be inactivated and no refunds will be made. -
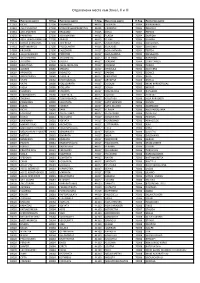
Print Sheet Remote1
Отдалечени места към Зона I, II и III П.Код Населено място П.Код Населено място П.Код Населено място П.Код Населено място 11361 KICELI 27100 KERAMIDIA 44015 LAGKADA 70004 KSEROKABOS 12351 AGIA VARVARA 27100 MONI FRAGKOPIDIMATOS 44015 LIKORRAXI 70004 PERVOLA 13561 AGII ANARGIRI 27100 TRAGANO 44015 OKSIA 70004 PEFKOS 13672 PARNITHA 27200 AGIA MARINA 44015 PLAGIA 70004 SKAFIDIA 13679 AGIA TRIADA PARNITHAS 27200 ANALICI 44015 PLIKATI 70004 STAFRIA 13679 KSENIA PARNITHAS 27200 ASTEREIKA 44015 PIRSOGIANNI 70004 SIKOLOGOS 14451 METAMORFOSI 27200 PALEOLANTHI 44015 XIONADES 70004 SINDONIA 14568 KRIONERI 27200 PALEOXORI 44017 AGIA VARVARA 70004 TERTSA 15342 AGIA PARASKEFI 27200 PERISTERI 44017 AGIA MARINA 70004 FAFLAGKOS 18010 AGIA MARINA 27300 AGIA MAFRA 44017 VEDERIKOS 70004 XONDROS 18010 AGKISTRI 27300 KALIVIA 44017 VERENIKI 70004 CARI FORADA 18010 EGINITISSA 28080 AGIOS NIKOLAOS 44017 VROSINA 70005 AVDOU 18010 ALONES 28080 GRIZATA 44017 VRISOULA 70005 ANO KERA 18010 APONISOS 28080 DIGALETO 44017 GARDIKI 70005 GONIES 18010 APOSPORIDES 28080 ZERVATA 44017 GKRIBOVO 70005 KERA 18010 VATHI 28080 KARAVOMILOS 44017 GRANITSA 70005 KRASIO 18010 VATHI 28080 KOULOURATA 44017 DIXOUNI 70005 MONI KARDIOTISSAS 18010 VIGLA 28080 POULATA 44017 DOVLA 70005 MOXOS 18010 VLAXIDES 28080 STAVERIS 44017 DOMOLESSA 70005 POTAMIES 18010 GIANNAKIDES 28080 TZANEKATA 44017 ZALOGO 70005 SFENDILI 18010 THERMA 28080 TSAKARISIANOS 44017 KALLITHEA 70006 AGIA PARASKEFI 18010 KANAKIDES 28080 XALIOTATA 44017 KATO VERENIKI 70006 AGNOS 18010 KLIMA 28080 XARAKTI 44017 KATO ZALOGO -

CRETE CHANIA AIRPORT – CHANIA Town and Surroundings
Service is available for flights between 1st April and 26th October CRETE CHANIA AIRPORT – CHANIA town and surroundings Shared door to door Service HOTELS ONLY! Chania town 14.70 EUR/passenger/way Daratso / Kato Galtatas / Ag. Apostoli/ Chrissi Akti / 15.80 EUR/passenger/way Kalamaki 16.90 EUR/passenger/way Platanias / Agia Marina / Stalos / Stavros / Akrotiri Meeting points: Crete Chania Airport: At the airport our meeting point is outside of the airport. Crete destinations, shared door-to-door service: In the city our driver will wait you at the hotel reception or the nearest possible place if the hotel cannot be reached by the coach. Shared door-to-door transfer to Crete destinations: Our city shuttle service is a shared service – we pick-up/drop-off passengers at different locations. Journey time: approximately 30-100 minutes, depending on the traffic. It is important that the booked transfer gets only confirmed if the correct destination/pick-up address is given (we accept addresses only within the served area). Bookings with not full or with no destination/pick up address will be inactivated and no refunds will be made. The service is available only to HOTELS. Contacts Call Center: 0036 30 622 89 19 E-mail: [email protected] (general information) For all questions about the transfer, please check FAQs on our website: http://ryanair.plusairportline.com/index.php?page=faq Our Travel Contract is available at the following link: http://ryanair.plusairportline.com/uploads/terms/terms_en.pdf Please note that we cannot guarantee bookings within 24 hours or for the following day.