Auditory Cortex ‐ 1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Five Topographically Organized Fields in the Somatosensory Cortex of the Flying Fox: Microelectrode Maps, Myeloarchitecture, and Cortical Modules
THE JOURNAL OF COMPARATIVE NEUROLOGY 317:1-30 (1992) Five Topographically Organized Fields in the Somatosensory Cortex of the Flying Fox: Microelectrode Maps, Myeloarchitecture, and Cortical Modules LEAH A. KRUBITZER AND MIKE B. CALFORD Vision, Touch and Hearing Research Centre, Department of Physiology and Pharmacology, The University of Queensland, Queensland, Australia 4072 ABSTRACT Five somatosensory fields were defined in the grey-headed flying fox by using microelec- trode mapping procedures. These fields are: the primary somatosensory area, SI or area 3b; a field caudal to area 3b, area 1/2; the second somatosensory area, SII; the parietal ventral area, PV; and the ventral somatosensory area, VS. A large number of closely spaced electrode penetrations recording multiunit activity revealed that each of these fields had a complete somatotopic representation. Microelectrode maps of somatosensory fields were related to architecture in cortex that had been flattened, cut parallel to the cortical surface, and stained for myelin. Receptive field size and some neural properties of individual fields were directly compared. Area 3b was the largest field identified and its topography was similar to that described in many other mammals. Neurons in 3b were highly responsive to cutaneous stimulation of peripheral body parts and had relatively small receptive fields. The myeloarchi- tecture revealed patches of dense myelination surrounded by thin zones of lightly myelinated cortex. Microelectrode recordings showed that myelin-dense and sparse zones in 3b were related to neurons that responded consistently or habituated to repetitive stimulation respectively. In cortex caudal to 3b, and protruding into 3b, a complete representation of the body surface adjacent to much of the caudal boundary of 3b was defined. -

'What' but Not 'Where' Auditory Processing Pathway
NeuroImage 82 (2013) 295–305 Contents lists available at SciVerse ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg Emotion modulates activity in the ‘what’ but not ‘where’ auditory processing pathway James H. Kryklywy d,e, Ewan A. Macpherson c,f, Steven G. Greening b,e, Derek G.V. Mitchell a,b,d,e,⁎ a Department of Psychiatry, University of Western Ontario, London, Ontario N6A 5A5, Canada b Department of Anatomy & Cell Biology, University of Western Ontario, London, Ontario N6A 5A5, Canada c National Centre for Audiology, University of Western Ontario, London, Ontario N6A 5A5, Canada d Graduate Program in Neuroscience, University of Western Ontario, London, Ontario N6A 5A5, Canada e Brain and Mind Institute, University of Western Ontario, London, Ontario N6A 5A5, Canada f School of Communication Sciences and Disorders, University of Western Ontario, London, Ontario N6A 5A5, Canada article info abstract Article history: Auditory cortices can be separated into dissociable processing pathways similar to those observed in the vi- Accepted 8 May 2013 sual domain. Emotional stimuli elicit enhanced neural activation within sensory cortices when compared to Available online 24 May 2013 neutral stimuli. This effect is particularly notable in the ventral visual stream. Little is known, however, about how emotion interacts with dorsal processing streams, and essentially nothing is known about the impact of Keywords: emotion on auditory stimulus localization. In the current study, we used fMRI in concert with individualized Auditory localization Emotion auditory virtual environments to investigate the effect of emotion during an auditory stimulus localization fi Auditory processing pathways task. Surprisingly, participants were signi cantly slower to localize emotional relative to neutral sounds. -

A Search for Factors Specifying Tonotopy Implicates DNER in Hair-Cell Development in the Chick's Cochlea☆
Developmental Biology 354 (2011) 221–231 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology A search for factors specifying tonotopy implicates DNER in hair-cell development in the chick's cochlea☆ Lukasz Kowalik 1, A.J. Hudspeth ⁎ Howard Hughes Medical Institute and Laboratory of Sensory Neuroscience, Campus box 314, The Rockefeller University, 1230 York Avenue, New York, NY 10065–6399, USA article info abstract Article history: The accurate perception of sound frequency by vertebrates relies upon the tuning of hair cells, which are Received for publication 1 March 2011 arranged along auditory organs according to frequency. This arrangement, which is termed a tonotopic Revised 29 March 2011 gradient, results from the coordination of many cellular and extracellular features. Seeking the mechanisms Accepted 31 March 2011 that orchestrate those features and govern the tonotopic gradient, we used expression microarrays to identify Available online 8 April 2011 genes differentially expressed between the high- and low-frequency cochlear regions of the chick (Gallus gallus). Of the three signaling systems that were represented extensively in the results, we focused on the Keywords: ζ Auditory system notch pathway and particularly on DNER, a putative notch ligand, and PTP , a receptor phosphatase that Hair bundle controls DNER trafficking. Immunohistochemistry confirmed that both proteins are expressed more strongly Planar cell polarity in hair cells at the cochlear apex than in those at the base. At the apical surface of each hair cell, the proteins Signaling display polarized, mutually exclusive localization patterns. Using morpholinos to decrease the expression of DNER or PTPζ as well as a retroviral vector to overexpress DNER, we observed disturbances of hair-bundle morphology and orientation. -

Anatomy of the Temporal Lobe
Hindawi Publishing Corporation Epilepsy Research and Treatment Volume 2012, Article ID 176157, 12 pages doi:10.1155/2012/176157 Review Article AnatomyoftheTemporalLobe J. A. Kiernan Department of Anatomy and Cell Biology, The University of Western Ontario, London, ON, Canada N6A 5C1 Correspondence should be addressed to J. A. Kiernan, [email protected] Received 6 October 2011; Accepted 3 December 2011 Academic Editor: Seyed M. Mirsattari Copyright © 2012 J. A. Kiernan. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Only primates have temporal lobes, which are largest in man, accommodating 17% of the cerebral cortex and including areas with auditory, olfactory, vestibular, visual and linguistic functions. The hippocampal formation, on the medial side of the lobe, includes the parahippocampal gyrus, subiculum, hippocampus, dentate gyrus, and associated white matter, notably the fimbria, whose fibres continue into the fornix. The hippocampus is an inrolled gyrus that bulges into the temporal horn of the lateral ventricle. Association fibres connect all parts of the cerebral cortex with the parahippocampal gyrus and subiculum, which in turn project to the dentate gyrus. The largest efferent projection of the subiculum and hippocampus is through the fornix to the hypothalamus. The choroid fissure, alongside the fimbria, separates the temporal lobe from the optic tract, hypothalamus and midbrain. The amygdala comprises several nuclei on the medial aspect of the temporal lobe, mostly anterior the hippocampus and indenting the tip of the temporal horn. The amygdala receives input from the olfactory bulb and from association cortex for other modalities of sensation. -

The Topographic Unsupervised Learning of Natural Sounds in the Auditory Cortex
The topographic unsupervised learning of natural sounds in the auditory cortex Hiroki Terashima Masato Okada The University of Tokyo / JSPS The University of Tokyo / RIKEN BSI Tokyo, Japan Tokyo, Japan [email protected] [email protected] Abstract The computational modelling of the primary auditory cortex (A1) has been less fruitful than that of the primary visual cortex (V1) due to the less organized prop- erties of A1. Greater disorder has recently been demonstrated for the tonotopy of A1 that has traditionally been considered to be as ordered as the retinotopy of V1. This disorder appears to be incongruous, given the uniformity of the neocortex; however, we hypothesized that both A1 and V1 would adopt an efficient coding strategy and that the disorder in A1 reflects natural sound statistics. To provide a computational model of the tonotopic disorder in A1, we used a model that was originally proposed for the smooth V1 map. In contrast to natural images, natural sounds exhibit distant correlations, which were learned and reflected in the disor- dered map. The auditory model predicted harmonic relationships among neigh- bouring A1 cells; furthermore, the same mechanism used to model V1 complex cells reproduced nonlinear responses similar to the pitch selectivity. These results contribute to the understanding of the sensory cortices of different modalities in a novel and integrated manner. 1 Introduction Despite the anatomical and functional similarities between the primary auditory cortex (A1) and the primary visual cortex (V1), the computational modelling of A1 has proven to be less fruitful than V1, primarily because the responses of A1 cells are more disorganized. -

Conserved Role of Sonic Hedgehog in Tonotopic Organization of the Avian Basilar Papilla and Mammalian Cochlea
Conserved role of Sonic Hedgehog in tonotopic organization of the avian basilar papilla and mammalian cochlea Eun Jin Sona,1, Ji-Hyun Mab,c,1, Harinarayana Ankamreddyb,c, Jeong-Oh Shinb, Jae Young Choia,c, Doris K. Wud, and Jinwoong Boka,b,c,2 Departments of aOtorhinolaryngology and bAnatomy, and cBK21 PLUS Project for Medical Science, Yonsei University College of Medicine, Seoul 120-752, South Korea; and dLaboratory of Molecular Biology, National Institute on Deafness and Other Communication Disorders, Bethesda, MD 20892 Edited by Clifford J. Tabin, Harvard Medical School, Boston, MA, and approved February 11, 2015 (received for review September 16, 2014) Sound frequency discrimination begins at the organ of Corti in several important questions remain about the development of mammals and the basilar papilla in birds. Both of these hearing the tonotopic organization of the cochlea: (i) What is the signaling organs are tonotopically organized such that sensory hair cells pathway(s) upstream of Bmp7 and RA? and (ii) are Bmp7 and at the basal (proximal) end respond to high frequency sound, RA also required for establishing the tonotopy in the mammalian whereas their counterparts at the apex (distal) respond to low inner ear? frequencies. Sonic hedgehog (Shh) secreted by the developing The Shh signaling pathway is of particular interest in cochlear notochord and floor plate is required for cochlear formation in tonotopic development for several reasons. First, the require- both species. In mice, the apical region of the developing cochlea, ment of Shh emanating from the floor plate and notochord closer to the ventral midline source of Shh, requires higher levels appears conserved between chicken and mouse (8–11). -

Brain Maps – the Sensory Homunculus
Brain Maps – The Sensory Homunculus Our brains are maps. This mapping results from the way connections in the brain are ordered and arranged. The ordering of neural pathways between different parts of the brain and those going to and from our muscles and sensory organs produces specific patterns on the brain surface. The patterns on the brain surface can be seen at various levels of organization. At the most general level, areas that control motor functions (muscle movement) map to the front-most areas of the cerebral cortex while areas that receive and process sensory information are more towards the back of the brain (Figure 1). Motor Areas Primary somatosensory area Primary visual area Sensory Areas Primary auditory area Figure 1. A diagram of the left side of the human cerebral cortex. The image on the left shows the major division between motor functions in the front part of the brain and sensory functions in the rear part of the brain. The image on the right further subdivides the sensory regions to show regions that receive input from somatosensory, auditory, and visual receptors. We then can divide these general maps of motor and sensory areas into regions with more specific functions. For example, the part of the cerebral cortex that receives visual input from the retina is in the very back of the brain (occipital lobe), auditory information from the ears comes to the side of the brain (temporal lobe), and sensory information from the skin is sent to the top of the brain (parietal lobe). But, we’re not done mapping the brain. -

Subdivisions of Auditory Cortex and Processing Streams in Primates
Colloquium Subdivisions of auditory cortex and processing streams in primates Jon H. Kaas*† and Troy A. Hackett‡ Departments of †Psychology and ‡Hearing and Speech Sciences, Vanderbilt University, Nashville, TN 37240 The auditory system of monkeys includes a large number of histochemical studies in chimpanzees and humans, and nonin- interconnected subcortical nuclei and cortical areas. At subcortical vasive functional studies in humans. levels, the structural components of the auditory system of mon- keys resemble those of nonprimates, but the organization at The Core Areas of Auditory Cortex cortical levels is different. In monkeys, the ventral nucleus of the Originally, auditory cortex of monkeys was thought to be orga- medial geniculate complex projects in parallel to a core of three nized much as in cats, with a single primary area, AI, in the primary-like auditory areas, AI, R, and RT, constituting the first cortex of the lower bank of the lateral sulcus and a second area, stage of cortical processing. These areas interconnect and project AII, deeper in the sulcus (e.g., ref. 3). This concept fits well with to the homotopic and other locations in the opposite cerebral the early view that auditory, somatosensory, and visual systems hemisphere and to a surrounding array of eight proposed belt all have two fields. However, we now know that primates have areas as a second stage of cortical processing. The belt areas in turn a number of sensory representations for each modality, and project in overlapping patterns to a lateral parabelt region with at several somatosensory and auditory fields can be considered least rostral and caudal subdivisions as a third stage of cortical primary or primary like in character. -

A Core Speech Circuit Between Primary Motor, Somatosensory, and Auditory Cortex
bioRxiv preprint doi: https://doi.org/10.1101/139550; this version posted May 18, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Speech core 1 A core speech circuit between primary motor, somatosensory, and auditory cortex: Evidence from connectivity and genetic descriptions * ^ Jeremy I Skipper and Uri Hasson * Experimental Psychology, University College London, UK ^ Center for Mind/Brain Sciences (CIMeC), University of Trento, Italy, and Center for Practical Wisdom, Dept. of Psychology, The University of Chicago Running title: Speech core Words count: 20,362 Address correspondence to: Jeremy I Skipper University College London Experimental Psychology 26 Bedford Way London WC1H OAP United Kingdom E-mail: [email protected] bioRxiv preprint doi: https://doi.org/10.1101/139550; this version posted May 18, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Speech core 2 Abstract What adaptations allow humans to produce and perceive speech so effortlessly? We show that speech is supported by a largely undocumented core of structural and functional connectivity between the central sulcus (CS or primary motor and somatosensory cortex) and the transverse temporal gyrus (TTG or primary auditory cortex). Anatomically, we show that CS and TTG cortical thickness covary across individuals and that they are connected by white matter tracts. -

A Core Speech Circuit Between Primary Motor, Somatosensory, and Auditory Cortex
bioRxiv preprint doi: https://doi.org/10.1101/139550; this version posted May 19, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Speech core 1 A core speech circuit between primary motor, somatosensory, and auditory cortex: Evidence from connectivity and genetic descriptions * ^ Jeremy I Skipper and Uri Hasson * Experimental Psychology, University College London, UK ^ Center for Mind/Brain Sciences (CIMeC), University of Trento, Italy, and Center for Practical Wisdom, Dept. of Psychology, The University of Chicago Running title: Speech core Words count: 20,362 Address correspondence to: Jeremy I Skipper University College London Experimental Psychology 26 Bedford Way London WC1H OAP United Kingdom E-mail: [email protected] bioRxiv preprint doi: https://doi.org/10.1101/139550; this version posted May 19, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Speech core 2 Abstract What adaptations allow humans to produce and perceive speech so effortlessly? We show that speech is supported by a largely undocumented core of structural and functional connectivity between the central sulcus (CS or primary motor and somatosensory cortex) and the transverse temporal gyrus (TTG or primary auditory cortex). Anatomically, we show that CS and TTG cortical thickness covary across individuals and that they are connected by white matter tracts. -
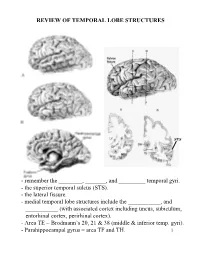
Review of Temporal Lobe Structures
REVIEW OF TEMPORAL LOBE STRUCTURES STS - remember the ________, _______, and _________ temporal gyri. - the superior temporal sulcus (STS). - the lateral fissure. - medial temporal lobe structures include the ___________, and ___________ (with associated cortex including uncus, subiculum, entorhinal cortex, perirhinal cortex). - Area TE = Brodmann’s 20, 21 & 38 (middle & inferior temp. gyri). - Parahippocampal gyrus = area TF and TH. 1 TEMPORAL LOBE FUNCTIONS Sensory Inputs to Temporal lobe: 1. ____________________________________________; 2. _________________________________________________. Temporal cortical regions and functional correlates: 1. Within lateral fissure (superior surface of lateral fissure): a) Heschel’s gyri (_____________________). b) posterior to Heschel’s gyri (_______________________). c) Planum temporale (secondary auditory cortex; Wernicke’s area - specialized in __________________________). 2 Temporal cortical regions and functional correlates (continued): 2. Superior temporal sulcus, middle and inferior temporal gyrus (area TE): _________________________________________ ____________. 3. Ventral/medial surface of temporal lobe (hippocampus and associated cortex): ______________________________. - the ventral/medial surface of the temporal lobe is also associated with the amygdala. Together with the surrounding ventral/medial temporal lobe, the amygdala is involved in __________________________________________. Hemispheric “specialization”: 1. Left hemisphere: a) ________________; b) ____________________________. -

Quantifying Attentional Modulation of Auditory-Evoked Cortical Responses from Single-Trial Electroencephalography
ORIGINAL RESEARCH ARTICLE published: 04 April 2013 HUMAN NEUROSCIENCE doi: 10.3389/fnhum.2013.00115 Quantifying attentional modulation of auditory-evoked cortical responses from single-trial electroencephalography Inyong Choi 1, Siddharth Rajaram 1, Lenny A. Varghese 1,2 and Barbara G. Shinn-Cunningham 1,2* 1 Center for Computational Neuroscience and Neural Technology, Boston University, Boston, MA, USA 2 Department of Biomedical Engineering, Boston University, Boston, MA, USA Edited by: Selective auditory attention is essential for human listeners to be able to communicate John J. Foxe, Albert Einstein College in multi-source environments. Selective attention is known to modulate the neural of Medicine, USA representation of the auditory scene, boosting the representation of a target sound Reviewed by: relative to the background, but the strength of this modulation, and the mechanisms Kimmo Alho, University of Helsinki, Finland contributing to it, are not well understood. Here, listeners performed a behavioral Sarah E. Donohue, Duke University, experiment demanding sustained, focused spatial auditory attention while we measured USA cortical responses using electroencephalography (EEG). We presented three concurrent *Correspondence: melodic streams; listeners were asked to attend and analyze the melodic contour of one Barbara G. Shinn-Cunningham, of the streams, randomly selected from trial to trial. In a control task, listeners heard the Auditory Neuroscience Laboratory, Center for Computational same sound mixtures, but performed the contour judgment task on a series of visual Neuroscience and Neural arrows, ignoring all auditory streams. We found that the cortical responses could be Technology, Boston University, fit as weighted sum of event-related potentials evoked by the stimulus onsets in the 677 Beacon St., Boston, competing streams.