New Insights on the Permian and Triassic Vertebrates from the Iberian Peninsula with Emphasis on the Pyrenean and Catalonian Basins
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Els Camins Antics I Les Vies Romanes a La Comarca D'osona
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Repositori Institucional de la Universitat Jaume I QUAD. PREH. ARQ. CAST. 28, 2010 Els camins antics i les vies romanes a la comarca d’Osona (Barcelona). Estat de la qüestió Carles Padrós* Resum En aquest estudi s’ha realitzat un estat de la qüestió de les vies i camins antics del que avui és la comarca d’Osona. Cal tenir present la importància del coneixement d’aquestes vies de comunicació a l’hora d’estudiar antics assentaments o zones d’hàbitat dins l’Ausetània ibèrica, romana o posterior, ja que sense aquestes no es podrien relacionar els assen- taments amb el seu entorn més proper o llunyà. En definitiva les vies i camins antics resulten de gran importància per entendre un espai des d’un anàlisis macro del territori. Abstract In this study we have upgraded the ancient roads and tracks of the Osona region. It must be taken into account the importance of knowing these channels of communication while studying ancient settlements or areas of habitat in the Ausetania Iberian period (or later), because without these, no settlement could be related to their immediate or distant environment.Therefore, ancient tracks and roads are of great importance for understanding an area from a macro analysis of the territory. profundes gorges, s’obre pas cap al Vallès Oriental desembocant en el riu Besòs. INTRODUCCIÓ PLANTEJAMENT I METODOLOGIA DELIMITACIÓ DEL MARC ESPACIAL I Aquest estudi pretén posar al dia el coneixe- TEMPORAL ment de les vies i camins d’època antiga a la comar- L’àrea geogràfica que centre aquest treball ca d’Osona (Fig. -

Revision Des Faunes De Vertebres Du Site De Proven Cheres-Sur-Meuse (Trias Terminal, Nord-Est De La France)
REVISION DES FAUNES DE VERTEBRES DU SITE DE PROVEN CHERES-SUR-MEUSE (TRIAS TERMINAL, NORD-EST DE LA FRANCE) par Gilles CUNY * SOMMAIRE Page Résumé, Abstract ..................... : . .. 103 Introduction ..................................................................... 103 Historique. 103 Révision des anciennes collections ................................................... 105 Muséum National d'Histoire Naturelle ................... 105 Université Pierre et Marie Curie. .. .. .. 106 Besançon . 120 Dijon. .. .. .. .. 121 Langres - Saint-Dizier. 121 Lyon ....................................................................... 123 Etude du matériel récent. 126 Discussion ...................................................................... 127 Conclusion ................................................................. :. 129 Remerciements. .. 130 Bibliographie . .. 130 Légendes des planches. 134 * Laboratoire de Paléontologie des Vertébrés, Université Pierre et Marie Curie - Boîte 106, 4 place Jussieu, 75252 PARIS cédex OS, France. Mots-clés: Trias, Rhétien, Poissons, Amphibiens, Reptiles Key-words: Triassic, Rhetian, Fishes, Amphibians, Reptiles PalaeoveT/ebra/a. Montpellier, 24 (1-2): 10H34. 6 fig .• 3 pl. (Reçu le 23 Février 1994. accepté le 15 Mai 1994. publié le 14 Juin 1995) -~----------------- - --.------- --------------- 500MHRES -- ------ - ------ -- --- ---- -------- ---.- A--- ..L -L ~ ~ 1.10 1.00 1 <6~::<:<=.: :~:~:::,'EE: J '_ 0.50 =...- ~ ::_-_____ -___ ---___ ~~:: ~-- __ .- _______ ~:: ----___ ~: ~-____ : ~ ~-. ~ ~ '_____ -

And Early Jurassic Sediments, and Patterns of the Triassic-Jurassic
and Early Jurassic sediments, and patterns of the Triassic-Jurassic PAUL E. OLSEN AND tetrapod transition HANS-DIETER SUES Introduction parent answer was that the supposed mass extinc- The Late Triassic-Early Jurassic boundary is fre- tions in the tetrapod record were largely an artifact quently cited as one of the thirteen or so episodes of incorrect or questionable biostratigraphic corre- of major extinctions that punctuate Phanerozoic his- lations. On reexamining the problem, we have come tory (Colbert 1958; Newell 1967; Hallam 1981; Raup to realize that the kinds of patterns revealed by look- and Sepkoski 1982, 1984). These times of apparent ing at the change in taxonomic composition through decimation stand out as one class of the great events time also profoundly depend on the taxonomic levels in the history of life. and the sampling intervals examined. We address Renewed interest in the pattern of mass ex- those problems in this chapter. We have now found tinctions through time has stimulated novel and com- that there does indeed appear to be some sort of prehensive attempts to relate these patterns to other extinction event, but it cannot be examined at the terrestrial and extraterrestrial phenomena (see usual coarse levels of resolution. It requires new fine- Chapter 24). The Triassic-Jurassic boundary takes scaled documentation of specific faunal and floral on special significance in this light. First, the faunal transitions. transitions have been cited as even greater in mag- Stratigraphic correlation of geographically dis- nitude than those of the Cretaceous or the Permian junct rocks and assemblages predetermines our per- (Colbert 1958; Hallam 1981; see also Chapter 24). -

Upper Triassic) of Guizhou, South China
Research Advances A New Discovery of Colobodus Agassiz, 1844 (Colobodontidae) from the Carnian (Upper Triassic) of Guizhou, South China LI Ji1,*, LUO Yongming2, WANG Yue1, XU Guangfu1, MA Zhiheng1,3 1 College of Resource and Environmental Engineering, Guizhou University, Guiyang 550025, China 2 Geological Survey of Guizhou, Guiyang 550004, China 3.Chongqing Institute of Geology and Mineral Resources, Chongqing 400042, China Corresponding author E-mail: [email protected] Objective Palaeoichthyology has been identified a research hotspot since abundant Triassic ichthyolite was discovered in Monte San Giorgio and South China. Critical review of the Colobodontidae reveals that this family has important research value. Furthermore, the family Perleididae Brough, 1931 and the probably paraphyletic ‘Perleidus group’ Gardiner & Schaeffer, 1989 have been implicitly regarded a synonym of the unsatisfactorily defined family Colobodontidae. Until 2002, Colobodontidae had been universally accepted as a significant taxon among all Triassic ichthyolite. However, the most colobodontids are probably confined to the Anisian and Ladinian in the Western Tethys. A well-preserved colobodontid discovered in Guizhou, South China, throws new light on its distribution and stratigraphic range. Methods A specimen was collected at the Wusha village of Xingyi City, Guizhou Province, South China and is preserved in the Geological Museum of Guizhou Province. The repair work was completed in a physical way in the laboratory of the Department of Geology, Guizhou University. Photographing the specimen was done by a 3D Microscope VHX-100k and drawings were made with reference to the digital photographs. Results This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. -

Exceptional Vertebrate Biotas from the Triassic of China, and the Expansion of Marine Ecosystems After the Permo-Triassic Mass Extinction
Earth-Science Reviews 125 (2013) 199–243 Contents lists available at ScienceDirect Earth-Science Reviews journal homepage: www.elsevier.com/locate/earscirev Exceptional vertebrate biotas from the Triassic of China, and the expansion of marine ecosystems after the Permo-Triassic mass extinction Michael J. Benton a,⁎, Qiyue Zhang b, Shixue Hu b, Zhong-Qiang Chen c, Wen Wen b, Jun Liu b, Jinyuan Huang b, Changyong Zhou b, Tao Xie b, Jinnan Tong c, Brian Choo d a School of Earth Sciences, University of Bristol, Bristol BS8 1RJ, UK b Chengdu Center of China Geological Survey, Chengdu 610081, China c State Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences (Wuhan), Wuhan 430074, China d Key Laboratory of Evolutionary Systematics of Vertebrates, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China article info abstract Article history: The Triassic was a time of turmoil, as life recovered from the most devastating of all mass extinctions, the Received 11 February 2013 Permo-Triassic event 252 million years ago. The Triassic marine rock succession of southwest China provides Accepted 31 May 2013 unique documentation of the recovery of marine life through a series of well dated, exceptionally preserved Available online 20 June 2013 fossil assemblages in the Daye, Guanling, Zhuganpo, and Xiaowa formations. New work shows the richness of the faunas of fishes and reptiles, and that recovery of vertebrate faunas was delayed by harsh environmental Keywords: conditions and then occurred rapidly in the Anisian. The key faunas of fishes and reptiles come from a limited Triassic Recovery area in eastern Yunnan and western Guizhou provinces, and these may be dated relative to shared strati- Reptile graphic units, and their palaeoenvironments reconstructed. -

From the Middle Triassic of Southern China
Bollettino della Società Paleontologica Italiana, 50 (2), 2011, 75-83. Modena, 31 ottobre 201175 A new species of the genus Perleidus (Actinopterygii: Perleidiformes) from the Middle Triassic of Southern China Cristina LOMBARDO, Zuo-Yu SUN, Andrea TINTORI, Da-Yong JIANG & Wei-Cheng HAO C. Lombardo, Dipartimento di Scienze della Terra “A. Desio”, Università degli Studi di Milano, I-20113, Milano, Italy; [email protected] Z.-Y. Sun, Department of Geology and Geological Museum, Peking University, Beijing 100871, People’s Republic of China; [email protected] A. Tintori, Dipartimento di Scienze della Terra “A. Desio”, Università degli Studi di Milano, I-20113, Milano, Italy; [email protected] D.-Y. Jiang, Department of Geology and Geological Museum, Peking University, Beijing 100871, People’s Republic of China; [email protected] W.-C. Hao, Department of Geology and Geological Museum, Peking University, Beijing 100871, People’s Republic of China; [email protected] KEY WORDS - Actinopterygians, Perleidiformes, Perleidus sinensis n. sp., Middle Triassic, Southern China, Tethys. ABSTRACT - Perleidus sinensis n. sp., a new species of “Subholostean” fossil fish of the order Perleidiformes is described herein on the basis of a single, well-preserved specimen collected from the Upper Member of the Guanling Formation (Pelsonian, Middle Anisian, Middle Triassic) outcropping near Luoping (Yunnan Province) in South China. The vertebrate assemblage yielded by these levels is proving to be of importance with regard to the marine Triassic -

Copyrighted Material
06_250317 part1-3.qxd 12/13/05 7:32 PM Page 15 Phylum Chordata Chordates are placed in the superphylum Deuterostomia. The possible rela- tionships of the chordates and deuterostomes to other metazoans are dis- cussed in Halanych (2004). He restricts the taxon of deuterostomes to the chordates and their proposed immediate sister group, a taxon comprising the hemichordates, echinoderms, and the wormlike Xenoturbella. The phylum Chordata has been used by most recent workers to encompass members of the subphyla Urochordata (tunicates or sea-squirts), Cephalochordata (lancelets), and Craniata (fishes, amphibians, reptiles, birds, and mammals). The Cephalochordata and Craniata form a mono- phyletic group (e.g., Cameron et al., 2000; Halanych, 2004). Much disagree- ment exists concerning the interrelationships and classification of the Chordata, and the inclusion of the urochordates as sister to the cephalochor- dates and craniates is not as broadly held as the sister-group relationship of cephalochordates and craniates (Halanych, 2004). Many excitingCOPYRIGHTED fossil finds in recent years MATERIAL reveal what the first fishes may have looked like, and these finds push the fossil record of fishes back into the early Cambrian, far further back than previously known. There is still much difference of opinion on the phylogenetic position of these new Cambrian species, and many new discoveries and changes in early fish systematics may be expected over the next decade. As noted by Halanych (2004), D.-G. (D.) Shu and collaborators have discovered fossil ascidians (e.g., Cheungkongella), cephalochordate-like yunnanozoans (Haikouella and Yunnanozoon), and jaw- less craniates (Myllokunmingia, and its junior synonym Haikouichthys) over the 15 06_250317 part1-3.qxd 12/13/05 7:32 PM Page 16 16 Fishes of the World last few years that push the origins of these three major taxa at least into the Lower Cambrian (approximately 530–540 million years ago). -

Distribution and Significance of Mesozoic Vertebrate Trace Fossil
Lockley et al.: Distribution and Significance of Mesozoic Vertebrate Trace Fossil DISTRIBUTION AND SIGNIFICANCE OF MESOZOIC VERTEBRATE TRACE FOSSILS IN DINOSAUR NATIONAL MONUMENT • MARTIN LOCKLEY +KELLY CONRAD +MARC PAQUETTE GEOLOGY DEPARTMENT + UNIVERSITY OF COLORADO DENVER • INTRODUCTION • - FIELD METHODS Dinosaur National Monument (DNM) During the summer of 1991 (end of year one encompasses a large area with surface exposures of and start of year two), reconaissance exploration for. Mesozoic rocks. Although only one isolated footprint vertebrate tracks at DNM was undertaken by Martin (from an unknown locality) had previously been Lockley, Kelly Conrad and Mark Paquette in June, discovered at DNM, there is a high potential for July and August. Following our initial success with preservation of fossil footprints in this area, as has tracksite discovery in the late Triassic Chinle/Popo been proven by discoveries in similar rocks in the Agie beds of the western part of DNM, we continued -region around DNM, and during research activity at to survey this unit throughout DNM. Tracks were DNM. As reported by Lockley et al. (1990), the found at all but the last locality. At all sites where purpose of this research is to seek, document and exposure permitted, we also measured the interpret vertebrate trace fossils in any of the ten stratigraphic position of trackbearing layers. potentially track-bearing Mesozoic stratigraphic units in which footprints might be discovered. We also began a collecting and replicating program that resulted in the acquisition of 27 original During the course of the first ·year (6/90-6/91) specimens · and replicas. Tracks were molded from the University of Colorado at Denver Research Group original specimens in the field using latex rubber, documented 11 localities where tracks were then replicated in the lab using plaster of Paris. -

Quaderns De Natura PEIXOS ACTINOPTERIGIS DEL
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Revistes Catalanes amb Accés Obert Quaderns de Vilaniu, 35: 55-87 (1999) ^^ Quaderns de Natura PEIXOS ACTINOPTERIGIS DEL TRIÀSIC MITJÀ DE LA PEDRA D'ALCOVER * per Joan Cartanyà Martí Paraules clau: actinopterigis, sistemàtica, Ladinià superior. Unitat d'Alcover. Resum: Els fòssils de les dolmicrites d'Alcover es coneixen des del 1963. La Unitat de les Dolomies Taulejades d'Alcover (pedra d'Alcover) té una potència de 70-80 m, i es troba en depressions interesculls. Estratigràficament es situa al Ladinià superior (Triàsic mitjà). Durant la deposició de la Unitat d'Alcover es van desenvolupar condicions anòxiques i hipersalines d'una forma periòdica. Les fàcies més abundants són constituïdes per capes de dolmicrites massives (d'entre 1 i 10 cm de gruix) i dolmicrites laminades molt fines. Localment apareixen capes ondulades amb plegaments i cabussaments. L'associació fòssil és al·lòctona i composta per grups florístics i faunístics com: plantes terrestres, celenterats, braquiòpodes, mol·luscs, artròpodes, equinoderms, peixos i rèptils, però la reputació d'aquests jaciments rau en el fet que els fòssils s'han conservat en dolmicrites i en la seva diversitat ictiològica. La fauna de peixos d'Alcover es caracteritza bàsicament per individus de talla mitjana. Els Saurichtiformes i els Perleidiformes són els ordres més comuns. Mitjançant aquest estudi els actinopterigis es classifiquen bàsicament a nivell genèric, i en alguns casos només a nivell de família. D'aquesta manera, i fins avui, han sigut prèviament indentificats 15 gèneres pertanyents a 11 famílies i nou ordres: Plycholepis, IBoreosomiis, Sauriclitliys, Colobodiis, Perleidus, IClenognaliclilltys, Peltoperleidiis, ICleithrolepididae indet., Pelíopleurus, Peripeltopleurus, IPlatysiagiim, Liiganoia, Eosemionotiis, Archaeosemionotus, Allolepidotiis, Eoeugnathiis, ICaturus, lOphiopsis, IPholidophoridae indet. -
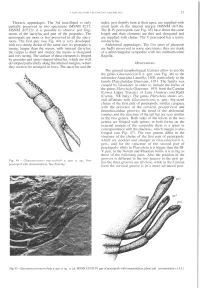
Thoracic Appendages. the 3Rd Maxilliped Is Only Partially
A NEW DECAPOD CRUSTACEAN ASSEMBLAGE 11 Thoracic appendages. The 3rd maxilliped is only index, just slightly bent at their apex, are supplied with partially preserved in two specimens (MSNB 8237; small teeth on the internal margin (MSNM i 10736). MSNM i 10732): it is possible to observe just frag- The II-IV pereiopods (see Fig. 45) have about the same ments of the dactylus and part of the propodus. The length and their elements are thin and elongated and pereiopods are more or less preserved in all the speci- are supplied with chelae. The V pereiopod has a termi- mens. The first pair (see Fig. 44) is very developed, nal dactylus. with two strong chelae of the same size; its propodus is Abdominal appendages. The five pairs of pleopods strong, longer than the merus, with internal dactylus; are badly preserved in some specimens; they are made the carpus is short and stocky; the merus is elongated of a subrectangular sympodite with two multiarticulate and very strong. The surface of these elements is fringed flagella. by granules and spine-shaped tubercles, which are well developed particularly along the internal margins, where Observations they seem to be arranged in rows. The dactylus and the The general morphological features allow to ascribe the genus Glaessnericaris n. gen. (see Fig. 46) to the infraorder Astacidea Latreille, 1803, particularly to the family Platychelidae Glaessner, 1931. The family was created by Glaessner in order to include the forms of the genus Platychela Glaessner, 1931 from the Carnian (Lower Upper Triassic) of Lunz (Austria) and Raibl (Carnia, NE Italy). -

Actinopterygian Fishes (Osteichthyes, Actinopterygii) from the Kalkschieferzone (Uppermost Ladinian) Near Meride (Canton Ticino, Southern Switzerland)
Actinopterygian fishes (Osteichthyes, Actinopterygii) from the Kalkschieferzone (Uppermost Ladinian) near Meride (Canton Ticino, Southern Switzerland) Autor(en): Bürgin, Toni Objekttyp: Article Zeitschrift: Eclogae Geologicae Helvetiae Band (Jahr): 88 (1995) Heft 3 PDF erstellt am: 10.10.2021 Persistenter Link: http://doi.org/10.5169/seals-167705 Nutzungsbedingungen Die ETH-Bibliothek ist Anbieterin der digitalisierten Zeitschriften. Sie besitzt keine Urheberrechte an den Inhalten der Zeitschriften. Die Rechte liegen in der Regel bei den Herausgebern. Die auf der Plattform e-periodica veröffentlichten Dokumente stehen für nicht-kommerzielle Zwecke in Lehre und Forschung sowie für die private Nutzung frei zur Verfügung. Einzelne Dateien oder Ausdrucke aus diesem Angebot können zusammen mit diesen Nutzungsbedingungen und den korrekten Herkunftsbezeichnungen weitergegeben werden. Das Veröffentlichen von Bildern in Print- und Online-Publikationen ist nur mit vorheriger Genehmigung der Rechteinhaber erlaubt. Die systematische Speicherung von Teilen des elektronischen Angebots auf anderen Servern bedarf ebenfalls des schriftlichen Einverständnisses der Rechteinhaber. Haftungsausschluss Alle Angaben erfolgen ohne Gewähr für Vollständigkeit oder Richtigkeit. Es wird keine Haftung übernommen für Schäden durch die Verwendung von Informationen aus diesem Online-Angebot oder durch das Fehlen von Informationen. Dies gilt auch für Inhalte Dritter, die über dieses Angebot zugänglich sind. Ein Dienst der ETH-Bibliothek ETH Zürich, Rämistrasse 101, 8092 Zürich, Schweiz, www.library.ethz.ch http://www.e-periodica.ch Eclogae geol. Helv. 88/3: 803-826 (1995) 0012-9402/95/030803-24 $1.50 + 0.20/0 Birkhäuser Verlag. Basel Actinopterygian fishes (Osteichthyes; Actinopterygii) from the Kalkschieferzone (Uppermost Ladinian) near Meride (Canton Ticino, Southern Switzerland) Tom Bürgin Key words: Kalkschieferzone. Middle Triassic. Monte San Giorgio, fossil Actinopterygii. -

Lab 5: Mollusks
Geos 223 Introductory Paleontology Spring 2006 Lab 5: Mollusks Name: Section: AIMS: This lab will introduce you to the eutrochozoan protostome phylum Mollusca. You will become familiar with the basic anatomy of the three mollusk groups which are most abundant in the fossil record: gastropods, bivalves, and cephalopods. Emphasis is placed on the various modes of life adopted by different members of each group, and how the form of the organism has been evolutionarily modified to suit each mode. You will also use a computer database to identify “mystery fossils”. By the end of this lab, you should have a good knowledge of the anatomy of the three most diverse groups of mollusks, an appreciation for how organismal form reflects function, and an understanding of how innovations in ecology and anatomy resulted in the evolutionary radiation of each group. INTRODUCTION: Mollusks are unsegmented protostomes with a trochophore larval stage during early development, and are one of the most diverse metazoan phyla. The basic mollusk body plan consists of a muscular foot, a visceral mass (containing the digestive tract and associated organs), a mantle cavity containing gills, a radula for feeding, and a calcareous shell protecting the visceral mass. The shell has a high preservation potential, and mollusks are common in the fossil record. There may be as many as ten classes of mollusks (depending on which text book you read). Each class has modified the basic body plan to some degree, allowing the group to radiate into different ecological niches. We will here focus on just three classes, which are common as fossils and exemplify the evolutionary diversification of mollusks.