Anatomical, Biomechanical and Physiological Loading During Human Endurance Performance at Selected Limb Cadences Via Triathlon
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

TE AWAMUTU COURIER, THURSDAY, MAY 5, 2011 Velodrome Proponents Approach
Te Awamutu COMPUTERS, SERVICE Courier NOTEBOOKS, SUPPORT APPLE PRODUCTS Published Tuesday & Thursday THURSDAY, MAY 5, 2011 NOW AT 407 Sloane Street, Te Awamutu Ph: 07 871 3837 Fax: 07 871 3807 CELEBRATING 100 YEARS AS YOUR COMMUNITY NEWSPAPER E-mail: [email protected] CIRCULATED FREE TO ALL HOUSEHOLDS THROUGHOUT TE AWAMUTU AND SURROUNDING DISTRICTS. EXTRA COPIES 40c. BRIEFLY Playful nature saves Stitch ‘Cybercrims’ capitalise Cybercriminals have been quick to pounce on Internet for Search and Rescue role users seeking information about the death of Osama bin Laden. BY GRANT JOHNSTON Within hours of the news A dog at Kihikihi pound who that US forces had killed the Al was due to be euthanised has Qaeda leader in Pakistan, instead been earmarked for a malware began popping up career in search and rescue. websites optimised to appear Stitch (as in ‘stitch in time’) is a on web searchers related to bin black and white, male, stock breed Laden’s death. and bull terrier cross. Scammers took advantage His breeding has mongrel of the phrase ‘Osama bin laden stamped all over it, but in nature dead’ being the most popular and manners he’s more like Prince Google search, luring Internet Charming. users to links supposedly He was tested on Monday for his containing photos, videos and suitability as an urban search and other information surrounding rescue (USAR) dog by training co- bin Laden’s death. ordinator for the upper North Many links have been Island region, Kirsti Kay. tainted with malware, infecting He came through the rigorous user’s computers when they testing with flying colours and are prompted to download a file video footage of his test will be sent away to the USAR national trainer or watch a video. -

The British Triathlon Federation
The British Triathlon Federation Minutes of the Annual General Meeting Held at 4:30 pm on Saturday 12 November 2011 at the Barceló Hinckley Island Hotel, Hinckley, Leicestershire Present: British Triathlon Triathlon England Dr Sarah Springman* OBE, President & Chair Derek Biggs1 David Rigby Zara Hyde Peters* OBE, Chief Executive Jem Lawson* Lawrence Green Ian Howard*, Independent Director Business Jane Vine John Lunt Ian Braid*, Director, Major & National Events Jane Wild John Petrides Ryan Wilkinson*, Independent Director, Finance Welsh Triathlon Howard Vine Chris Kitchen*, Director, Performance Dr Dean Hardie*1 Martin Harris Malcolm Westwood*, Director, Age Group Teams Steve Butler Dave Bellingham Neill Craigie*, Director, Marketing & Communications Tom Roberts Graeme Maw Tom Chambers*, triathlonscotland representative Bethan Fowler Richard Fuller Triathlonscotland Sue Fuller Geoff Earl1 Nina Foord Paul McGreal Gemma Simpson Mark Barfield, Director of Development and Maisie Bancewicz English Operations Dechlan Wilson Helen Wyeth, Director of Operations Judith Brand (Minutes) 1Voting Member, with names of accompanying Delegates underlined * BTF Board Member Completed by JB on 22 Nov 2011, reviewed by SMS on 3 Dec 2011, distributed to the Board and Delegates on 5 Dec 2011 and agreed on 12 Dec 2011 1 Opening Remarks The President (SMS) welcomed those present to the fifth Annual General Meeting of the British Triathlon Federation. The meeting observed a minute of silence in acknowledgement of Remembrance Day. SMS expressed thanks to John Lunt, who had kindly agreed to deliver a short presentation prior to the formal business of the meeting. SMS was pleased to welcome Derek Biggs, the newly elected President of Triathlon England. Apologies Paul Groves 2 President’s Report (see Appendix 1) The President gave her report of the activities of the Federation since the previous Annual General Meeting. -

2008 ITU Congress and Federations Forum
EUROPEAN TRIATHLON UNION ETU EVENT STATUS: 2001 – 2016 1. INTRODUCTION: The purpose of this document is to provide details about the status of the past and current ETU Events, including strengths and weaknesses. Recommendations will follow each section to provide an opportunity for further discussion with ITU and at the various levels at ETU. ETU main goals on events are: a) Increase the general participation of triathlon and all related multisport in all age-group categories within Europe; b) Provide opportunities for young athletes to access the sport; c) Keep gender balance on all level of competitions; d) Provide a pathway to grow from youth athlete to elite athletes competing at the Olympic Games. 2. GENERAL: A thorough review of the Competition System is required due to: a) the growing number of ITU events in Europe; b) the increased number of participants in all categories; c) the growing demands of Paratriathlon being integrated in the sport; d) the increased number of private international event organising companies wanting to align with our system. 3. ETU TRIATHLON EUROPEAN CHAMPIONSHIPS: is the main event of ETU. The event package consist of the following races: elite Olympic Distance Triathlon, the junior Sprint Triathlon event, the Age Group OD and sprint Triathlon, the elite and junior mixed relay, an opening ceremony and the organisation of the ETU Annual General Assembly. ETU is asking to pay an event fee and prize money for both the elite individual event as the elite mixed relay. Having the event live broadcasting is recommended but not mandatory. This means that the LOC has to invest a substantial amount into this organisation. -

Itu World Triathlon Series | Auckland | Sandiego | Yokohama | Madrid | Kitzbühel | Hamburg | Stockholm | London
2013 SERIES GUIDE ITU WORLd tRIATHLON SERIES | AUCKLAND | SAN DIEGO | YOKOHAMA | MADRID | KITZBÜHEL | HAMBURG | STOCKHOLM | LONDON ITU WORLD TRIATHLON SERIES | 2013 SERIES GUIDE 2 MEDIA CONTACTS ERIN GREENE MORGAN INGLIS Media Manager, ITU Communications Senior Producer, TV & Broadcast, ITU [email protected] [email protected] Office: + 34 915 421 855 Office: +1 604 904 9248 Mob: +34 645 216 509 Mobile: +1 604 250 4091 CARSTEN RICHTER OLIVER SCHIEK Upsolut Senior Director - TV Rights Upsolut Senior Director - TV Production [email protected] [email protected] Direct: +49 40 88 00 - 73 Direct: +49 40 88 18 00 - 48 Mobile: +49 170 56 39 008 Mobile: +49 170 34 29 886 ITU MEDIA CENTRE | MEDIA.TRIATHLON.ORG ITU’s Online Media Centre has been produced to provide a portal for media to quickly gather all relevant information about ITU, its events and athletes. Media Centre services include: • Latest ITU news and press releases • Up-to-date results, rankings and race statistics • Comprehensive athlete profile database • Rights-free high-resolution photos from all major events • Full audio from athlete interviews • Access to broadcast quality race video highlights For more information, or to register for a Media Centre account, visit media.triathlon.org. 3 2013 SERIES GUIDE | Itu WORLD TRIATHLON SERIES TABLE OF CONTENTS WELCOME TO THE SERIES Welcome from ITU President ..................................................... 04 Series Overview ������������������������������������������������������������������������� 05 -

2018 Wts Media Guide
WORLD TRIATHLON SERIES 1 2018 SERIES MEDIA GUIDE 2018 WTS MEDIA GUIDE ITU WORLD TRIATHLON SERIES | ABU DHABI | BERMUDA | YOKOHAMA | NOTTINGHAM / LEEDS | HAMBURG | EDMONTON | MONTREAL | GOLD COAST 2 WORLD TRIATHLON SERIES 2018 SERIES MEDIA GUIDE ITU MEDIA CONTACTS OLALLA CERNUDA FERGUS MURRAY Head of ITU Communications ITU TV & Broadcast [email protected] [email protected] Office: + 34 915 421 855 Office: +1 604 904 9248 Mob: +34 645 216 509 CHELSEA WHITE ADAM MASON Communication Specialist, ITU Communications TV Sales – Director Summer Sports, InFront [email protected] [email protected] Mob: +1 231 590 4026 Mob: +49 79 755 4292 DOUG GRAY Media Delegate, ITU Communications [email protected] Mob: +44 7583 620749 ITU MEDIA CENTRE | MEDIA.TRIATHLON.ORG ITU’s Online Media Centre has been produced to provide a portal for media to quickly gather all relevant information about ITU, its events and athletes. Media Centre services include: • Latest ITU news and press releases • Up-to-date results, rankings and race statistics • Comprehensive athlete profile database • Rights-free high-resolution photos from all major events • Full audio from athlete interviews • Access to broadcast quality race video highlights For more information, or to register for a Media Centre account, visit media.triathlon.org WORLD TRIATHLON SERIES 3 2018 SERIES MEDIA GUIDE TABLE OF CONTENTS WELCOME TO THE ITU TRIATHLON WORLD SERIES 4 SERIES OVERVIEW 5 ITU TRIATHLON HISTORY 6 THE BASICS 6 PARTNERSHIPS – THE COLLABORATIVE FORCES -
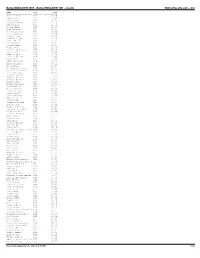
Bolderboulder 2005 - Bolderboulder 10K - Results Onlineraceresults.Com
BolderBOULDER 2005 - BolderBOULDER 10K - results OnlineRaceResults.com NAME DIV TIME ---------------------- ------- ----------- Michael Aish M28 30:29 Jesus Solis M21 30:45 Nelson Laux M26 30:58 Kristian Agnew M32 31:10 Art Seimers M32 31:51 Joshua Glaab M22 31:56 Paul DiGrappa M24 32:14 Aaron Carrizales M27 32:23 Greg Augspurger M27 32:26 Colby Wissel M20 32:36 Luke Garringer M22 32:39 John McGuire M18 32:42 Kris Gemmell M27 32:44 Jason Robbie M28 32:47 Jordan Jones M23 32:51 Carl David Kinney M23 32:51 Scott Goff M28 32:55 Adam Bergquist M26 32:59 trent r morrell M35 33:02 Peter Vail M30 33:06 JOHN HONERKAMP M29 33:10 Bucky Schafer M23 33:12 Jason Hill M26 33:15 Avi Bershof Kramer M23 33:17 Seth James DeMoor M19 33:20 Tate Behning M23 33:22 Brandon Jessop M26 33:23 Gregory Winter M26 33:25 Chester G Kurtz M30 33:27 Aaron Clark M18 33:28 Kevin Gallagher M25 33:30 Dan Ferguson M23 33:34 James Johnson M36 33:38 Drew Tonniges M21 33:41 Peter Remien M25 33:45 Lance Denning M43 33:48 Matt Hill M24 33:51 Jason Holt M18 33:54 David Liebowitz M28 33:57 John Peeters M26 34:01 Humberto Zelaya M30 34:05 Craig A. Greenslit M35 34:08 Galen Burrell M25 34:09 Darren De Reuck M40 34:11 Grant Scott M22 34:12 Mike Callor M26 34:14 Ryan Price M27 34:15 Cameron Widoff M35 34:16 John Tribbia M23 34:18 Rob Gilbert M39 34:19 Matthew Douglas Kascak M24 34:21 J.D. -

Relatório De Atividades 2019
RELATÓRIO DE ATIVIDADES 2019 Acredito que os bons resultados alcançados ao longo do ano de 2019 só existem porque temos uma equipe dedicada e comprometida, tanto nas atividades meio quanto nas atividades m. Dessa forma, entendo que as medalhas conquistadas em 2019 e o crescente desempenho dos nossos atletas são resultado do trabalho árduo – de cada atleta individualmente, é claro, mas de todos os prossionais que se dedicam tanto para que estejamos sempre próximos à excelência. Agradeço a cada um que se dedicou e acreditou no nosso esporte e na nossa Confederação. Ernesto Pitanga, Presidente da Confederação Brasileira de Triathlon A Confederação Brasileira de Triathlon está entre as Em 2019 superamos uma a uma as diculdades que confederações que possuem um plano de gestão se apresentaram, contando sempre com o nosso baseado nas necessidades do mundo moderno. time esportista, que possui o DNA de alto Calçamos nossos valores na ética, transparência, rendimento e compreende todo o processo de resiliência e muitos outros que se misturam com os formação de um atleta, passando pelos amadores, valores dos atletas que representamos. categoria de base e atletas paralímpicos. Temos orgulho em apresentar nosso relatório, Sabemos que o nosso dever é elevar o nosso esporte sabendo que conquistamos mais um degrau em a outro patamar, trazendo para casa a tão sonhada direção ao nosso objetivo – que, é importante medalha olímpica. armar, ainda se encontra distante do que pretendemos: a excelência é a nossa meta. Armando Barcellos, Vice-presidente da Confederação Brasileira de Triathlon Prezados, O ano de 2019 foi marcado pelos bons resultados – não me rero somente às medalhas dos Jogos Pan-Americanos de Lima e a todas as outras conquistas que nossos atletas trouxeram para casa e que tanto nos orgulham. -

2010 XTERRA Worlds Guide ROUGH:2007 XTERRA Maui Press
2010 PRESS GUIDE XTERRA WORLD CHAMPIONSHIP SPONSORS The XTERRA World Championship is presented by Paul Mitchell, Degree Men Adventure, Maui Visitors Bureau, Makena Beach & Golf Resort, and Hawaiian Airlines. Sponsors include GU, Gatorade, Zorrel, Kona Brewing Company, Hawaii Tourism Authority, XTERRA.TV, Hawaii Water, and the XTERRA Alliance - Gear, Footwear, Fitness, Wetsuits, and Cycling. THE 15TH ANNUAL XTERRA WORLD CHAMPIONSHIP . The XTERRA World Championship is the final stop on the XTERRA World Tour - a national and international series of 100+ qualifying events held in Austria, Brazil, Canada, Czech Republic, France, Germany, Italy, Japan, Mexico, New Zealand, Portugal, Saipan, South Africa, Switzerland, and the United States. The course, considered XTERRA’s toughest, consists of a 1.5-kilometer rough water swim, a grueling 32km mountain bike on the lower slopes of Haleakala, and an 11km trail run. The field is limited to 550 competitors, including 75 pros, who represent the best off-road multisport athletes on the planet. They come from more than 30 countries & compete for one of the richest purses in multisport at $100,000. The award-winning TEAM TV crew will be on location to televise all the action for a one-hour sports special that will air across the U.S. via national syndication starting in January of 2011. You can watch last year’s race show now at www.XTERRA.tv and get live text updates from Maui on raceday at www.xterramaui.com. TABLE OF CONTENTS Media Information . .4 Schedule of Events . .5 How to Watch Guide for Spectators . .6 World Championship Quick Facts . .7 Maui No Ka Oi translates to XTERRA Makena Beach Trail Runs . -

Statistics Investigation
Statistics Investigation Planning Introduction I will investigate performance in the Olympic triathlon event using data gathered from the Olympic results website. The first Olympic triathlon was held at the Summer 2000 Olympics so data should be available for three Olympics (2000, 2004 and 2008). Men and women compete in separate events. I will use a census of the individual years and genders because in Autograph I can easily manipulate large data sets (there are typically 50 competitors in each event). Autograph will enable me to compare statistics for each data set, using box and whisker diagrams showing extreme values, quartiles and the inter-quartile range. I will also create scatter graphs of one data parameter against another so that I can see if there is any obvious relationship between them. Autograph can also easily fit trend lines (to enable predictions to be made) and calculate the Spearman’s ranking coefficient. A strong correlation could indicate, for instance, that the results in one sport were very likely to influence the overall results whilst a weak correlation would indicate that an athlete could do well or badly in that sport and still be successful overall. I am hoping to find what kind of athlete does best in a triathlon: must they be very good in all three sports, or are some sports more critical than others? I will also investigate the differences between men’s and women’s times to see whether the same patterns are seen in both and whether conclusions from the men’s race are equally valid for the women’s competition. -

Itu World Cup Series History & Stats
ITU WORLD CUP SERIES HISTORY & STATS Last Updated: September 16th , 2014 All-Time Career World Cup Series Events winners *denotes still active Rank Men WC Wins Rank Women WC Wins 1º Brad Beven (AUS) 17 1º Vanessa Fernandes (POR) 20 2º Javier Gomez* (ESP) 14 2º Emma Carney (AUS) 19 3º Simon Whitfield (CAN) 12 3º Carol Montgomery (CAN) 15 4º Hamish Carter (NZL) 11 4º Loretta Harrop (AUS) 12 5º Miles Stewart (AUS) 8 4º Michellie Jones (AUS) 12 6º Simon Lessing (GBR) 7 6º Siri Lindley (USA) 11 6º Greg Welch (AUS) 7 7º Emma Snowsill (AUS) 10 6º Courtney Atkinson (AUS) 7 8º Rina (Bradshaw) Hill (AUS) 8 6º Brad Kahlefeldt (AUS) 7 9º Anja Dittmer* (GER) 7 10º Craig Walton (AUS) 6 9º Samantha Warriner (NZL) 7 Greg Bennett (USA) (He was 10º Australian) 6 9º Karen Smyers (USA) 7 10º Tim Don (GBR) 6 12º Melissa Mantak (USA) 5 13º Rasmus Henning (DEN) 5 12º Nicola Spirig* (SUI) 5 13º Bevan Docherty (NZL) 5 14º Laura (Reback) Bennett* (USA) 4 13º Kris Gemmell (NZL) 5 14º Jackie Gallagher (AUS) 4 16º Dmitriy Gaag (KAZ) 4 14º Barbara Lindquist (USA) 4 16º Andrew Johns (GBR) 4 14º Annabel Luxford (AUS) 4 16º Ivan Rana* (ESP) 4 14º Emma Moffatt* (AUS) 4 16º Peter Robertson (AUS) 4 14º Jenny Rose (NZL) 4 16º Andrew MacMartin (CAN) 4 14º Ai Ueda* (JPN) 4 16º Chris McCormack (AUS) 4 21º Jill Savege (CAN) 3 16º Mike Pigg (USA) 4 21º Sheila Taormina (USA) 3 16º Hunter Kemper* (USA) 4 21º Sabine Westhoff (GER) 3 24º Reinaldo Colucci* (BRA) 2 21º Andrea Whitcombe (GBR) 3 24 Mario Mola* (ESP) 2 21º Gwen Jorgensen* (USA) 3 24 Sven Riederer* (SUI) 2 21º Katie Hursey* -

Chasing 2:40 Despite Deeper Pro Fields
Chasing Despite deeper pro fi elds and constant advancements in bike 2:40 and run technol ogy, the best Ironman athletes in the world are still chasing after the elusive run record on the Big Island—a record that has stood for 23 years (and counting). BY KIM McDONALD JEFF CLARK 46 JANUARY/FEBRUARY 2013 47 foundation serious challenger, except Marino Vanhoenacker, who had earned an [open marathon] eight-minute advantage over the fi eld with an all-or-nothing solo potential in Kona.” According to six-time Ironman breakaway. Jacobs quickly chipped away at his advantage o the bike, Asked why no world champ Dave Scott, the and the Australian took the lead for good at mile 15 in the run. one has run faster key to effi cient running tech- Realizing he had a big lead once past Vanhoenacker, Jacobs dialed than he and Allen nique is a strong core—and he back his pace to avoid cramping and risking the win. “I walked did back in 1989, doesn’t mean a six-pack. through the ice stations of most aid stations and slowed down to Scott said he drink,” he said. “I knew I had a four- to fi ve-minute lead out of the doesn’t have a good Energy Lab over Andreas, whom I outran last year when I felt worse, answer. What’s so that gave me confi dence to relax and focus on making it home in especially puzzling one piece. I couldn’t have gone much faster. I had worked very hard to him is that many on the bike, and I was a little fl at on the run.” of the athletes who show up in Kona year after year are The Perfect Storm faster runners than Why no one has been able to run faster than Allen or Scott did in he and Allen ever Kona in 1989 is one of triathlon’s enduring mysteries, especially were, at least over considering all of the improvements that have been made over the shorter distances. -

Curriculum Vitea
CV – Jul Clonen – april 2015 Curriculum vitea Naam: CLONEN Voornaam: Julien C.F. à Calling name: Jul or Julian Place of living: Home address: Hasseltsestraat 45 – 3290 DIEST – BELGIUM Private Email address: [email protected] Day of birth: 02/05/1963 Place of Birth: Geel (Belgium) Study: § Physical education, teacher of Sports (1989). § Belgian Olympic Academy (1998) – management of HP federations and structures § Sport management (2007) Current occupation: * Team manager – Head coach of the Belgian Triathlon Federation Responsible for the international program of the elite athletes (all WTS and several WC’s) and development of the U23 and juniors into Belgium. * Member of the ITU coaches Commission * Owner of the THE SPORTAMBASSADORS Professional targets à Contribute to the growth and development of triathlon in Belgium, with the focus on high performance and coach education programs. à Share my knowledge and experiences with other coaches in Belgium and worldwide. Contribute to the personal development of the colleagues I work with on a daily basis. (Visiting and talking with teams, Clubs, Coaches, athletes, parents, …) à Enhance my personal expertise in high performance coaching, coach development and high performance management. 2015-2013: Combination part time Team manager / Head Coach into the triathlon Federation and part time Director of education High Performance Cycling into the Belgian Cycling Federation (Flemish Part) 2005-2013: Expert of the high performance sport in Flanders (Belgium). (8 years) A function in the government