Sperm Quality and Quantity Evolve Through Different Selective Processes in the Phasianidae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Big Year Ebook Free Download
THE BIG YEAR PDF, EPUB, EBOOK Mark Obmascik | 320 pages | 08 Dec 2011 | Transworld Publishers Ltd | 9780857500694 | English | London, United Kingdom The Big Year PDF Book Pete Shackelford Steve Darling Archived from the original on January 26, In , Nicole Koeltzow reached the species milestone on July 1, while in August Gaylee and Richard Dean became the first birders to reach species in consecutive years. Highway runs along the California Coast. Crazy Credits. By Noah Strycker July 26, Stu is hiking with his toddler grandson already enamored by birds in the Rockies. Mary Swit Calum Worthy Miller Greg Miller It also replaces Jack Black's narration of the story with a new narration by John Cleese who also receives a credit in the opening title sequence. Narrator voice Jack Black Brad is a skilled birder who can identify nearly any species solely by sound. Category:Birds and humans Zoomusicology. Added to Watchlist. Retrieved January 25, Get Audubon in Your Inbox Let us send you the latest in bird and conservation news. Retrieved Jessica Steve Martin Paul Lavigne Heather Osborne Share this page:. Birds class : Aves. Yukon News. Tony Cindy Busby Wheel of Fortune Underscore. Visit our What to Watch page. Caprimulgiformes nightjars and relatives Steatornithiformes Podargiformes Apodiformes swifts and hummingbirds. In , an unprecedented four birders attempted simultaneous ABA Area big years. Steve's character provides fatherly guidance and support that helps Jack Black's character move forward with his life and relationships. The company is in the middle of complicated negotiations to merge with a competitor, so his two anointed successors keep calling him back to New York for important meetings; to some extent he is a prisoner of his own success. -

Notes on Game & Game Shooting
RSITY OF CALIFORNIA LIBRARY OF THE UNIVERSITY OF CALIFORNIA LI RSITY OF CALIFORNIA LIBRARY OF THE UNIVERSITY OF CALIFORNIA LI 1 CHIFi IE UNIVERSITY OF CALIFORKU LIBRARY OF HE UltlVERSITY <5V HE UNIVERSITY OF CALIFORNIA LIBRARY OF THE UNIVERSITY OF CALIFORN NOTES ON GAME & GAME SHOOTING. MISCELLANEOUS OBSERVATIONS ON BIRDS AND ANIMALS, AND ON THE SPORT THEY AFFORD FOR THE GUN IN GREAT BRITAIN. ILLUSTRATED. BY J. J.jMANLEY, M.A., " AUTHOR OF NOTES" ON FISH AND FISHING/' ETC. LONDON: THE BAZAAR" OFFICE, 170, STRAND, W.C. LONDON : PRINTED BY ALFRED BRADLEY, 170, STRAND, W.C. OK PREFACE. THESE Notes, good reader or, as I should prefer to address you, brother sportsman were, in substance, contributed from time to time to the columns of the Country ; and are now, with some additions and cor- rections, presented in book form. They do not aspire to be considered as a complete natural history of the objects of the home sportsman's pursuit, nor as an exhaustive treatise on the science and practice of shooting. The small size of the volume precludes such an ambitious idea. They hardly aspire to be more than miscellaneous notes and jottings, the of and observation results, mainly, my own experience ; and if sportsmen generally, young and old especially the former take any pleasure in their perusal, one object, at least, of their publication will be attained. But though the volume is of a somewhat discursive character, each chapter is written on a more or less iv Preface. definite plan, the nomenclature, the natural history, the method of pursuit, and the gastronomic merits of each bird and beast of sport being discussed in order. -

Monitoring and Population Assessment of Baer's Pochard In
Monitoring An assessment of the wintering popula4on of Baer’s Pochard in central Myanmar Thiri Dae We Aung, Thet Zaw Naing, Saw Moses, Lay Win, Aung Myin Tun, Thiri Sandar Zaw and Simba Chan May 2016 Biodiversity And Nature Conservation Association Page May 2016 Submitted To:Oriental Bird Club P.O.Box 324, Bedford, MK42 0WG, United Kingdom. Submitted By: Thiri Dae We Aung1, Thet Zaw Naing2, Saw Moses3, Lay Win4, Aung Myin Tun5, Thiri Sandar Zaw6, Simba Chan7 1 Biodiversity And Nature Conservation Association, Myanmar 2 Wildlife Conservation Society, Myanmar 3 4 5 6 Biodiversity And Nature Conservation Association, Myanmar 7 BirdLife International, Tokyo, Japan To obtain copies of this report contact: Biodiversity And Nature Conservation Association, No.943(A), 2nd floor, Kyeikwine Pagoda Road, Mayangone Township, Yangon, Myanmar. [email protected] Front Photo Caption: Sighting Baer’s Pochard at Pyu Lake (photo by: Simba Chan) Suggested citation: Aung, T.D, T.Z. Naing, S. Moses, L. Win, A.M. Tun, T.S. Zaw & S. Chan. 2016. An assessment of the wintering population of Baer’s Pochard in central Myanmar. Unpublished report, Biodiversity And Nature Conservation Association: ?? pp. Biodiversity And Nature Conservation Association Page Table of Contents ABSTRACT..............................................................................................................................................3 INTRODUCTION.....................................................................................................................................4 -

Phylogenetic Relationships of the Phasianidae Reveals Possible Non-Pheasant Taxa
Journal of Heredity 2003:94(6):472–489 Ó 2003 The American Genetic Association DOI: 10.1093/jhered/esg092 Phylogenetic Relationships of the Phasianidae Reveals Possible Non-Pheasant Taxa K. L. BUSH AND C. STROBECK From the Department of Biological Sciences, University of Alberta–Edmonton, Edmonton, Alberta T6G 2E9, Canada. Address correspondence to Krista Bush at the address above, or e-mail: [email protected]. Abstract The phylogenetic relationships of 21 pheasant and 6 non-pheasant species were determined using nucleotide sequences from the mitochondrial cytochrome b gene. Maximum parsimony and maximum likelihood analysis were used to try to resolve the phylogenetic relationships within Phasianidae. Both the degree of resolution and strength of support are improved over previous studies due to the testing of a number of species from multiple pheasant genera, but several major ambiguities persist. Polyplectron bicalcaratum (grey peacock pheasant) is shown not to be a pheasant. Alternatively, it appears ancestral to either the partridges or peafowl. Pucrasia macrolopha macrolopha (koklass) and Gallus gallus (red jungle fowl) both emerge as non-pheasant genera. Monophyly of the pheasant group is challenged if Pucrasia macrolopha macrolopha and Gallus gallus are considered to be pheasants. The placement of Catreus wallichii (cheer) within the pheasants also remains undetermined, as does the cause for the great sequence divergence in Chrysolophus pictus obscurus (black-throated golden). These results suggest that alterations in taxonomic -

Sperm Storage in Caecilian Amphibians Susanne Kuehnel1 and Alexander Kupfer1,2*
Kuehnel and Kupfer Frontiers in Zoology 2012, 9:12 http://www.frontiersinzoology.com/content/9/1/12 SHORT REPORT Open Access Sperm storage in caecilian amphibians Susanne Kuehnel1 and Alexander Kupfer1,2* Abstract Background: Female sperm storage has evolved independently multiple times among vertebrates to control reproduction in response to the environment. In internally fertilising amphibians, female salamanders store sperm in cloacal spermathecae, whereas among anurans sperm storage in oviducts is known only in tailed frogs. Facilitated through extensive field sampling following historical observations we tested for sperm storing structures in the female urogenital tract of fossorial, tropical caecilian amphibians. Findings: In the oviparous Ichthyophis cf. kohtaoensis, aggregated sperm were present in a distinct region of the posterior oviduct but not in the cloaca in six out of seven vitellogenic females prior to oviposition. Spermatozoa were found most abundantly between the mucosal folds. In relation to the reproductive status decreased amounts of sperm were present in gravid females compared to pre-ovulatory females. Sperm were absent in females past oviposition. Conclusions: Our findings indicate short-term oviductal sperm storage in the oviparous Ichthyophis cf. kohtaoensis. We assume that in female caecilians exhibiting high levels of parental investment sperm storage has evolved in order to optimally coordinate reproductive events and to increase fitness. Keywords: Reproduction, Sperm storage, Amphibians, Caecilians Background sperm storage has also evolved in the third group of ex- Animal reproductive strategies include variable modes of tant amphibians, the limbless caecilians [5,6]. sperm transfer, fertilization, and type of offspring devel- Caecilians perform internal fertilization with the aid opment. -
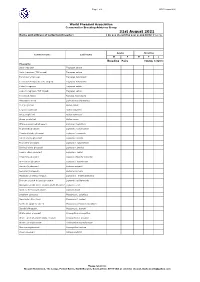
31St August 2021 Name and Address of Collection/Breeder: Do You Closed Ring Your Young Birds? Yes / No
Page 1 of 3 WPA Census 2021 World Pheasant Association Conservation Breeding Advisory Group 31st August 2021 Name and address of collection/breeder: Do you closed ring your young birds? Yes / No Adults Juveniles Common name Latin name M F M F ? Breeding Pairs YOUNG 12 MTH+ Pheasants Satyr tragopan Tragopan satyra Satyr tragopan (TRS ringed) Tragopan satyra Temminck's tragopan Tragopan temminckii Temminck's tragopan (TRT ringed) Tragopan temminckii Cabot's tragopan Tragopan caboti Cabot's tragopan (TRT ringed) Tragopan caboti Koklass pheasant Pucrasia macrolopha Himalayan monal Lophophorus impeyanus Red junglefowl Gallus gallus Ceylon junglefowl Gallus lafayettei Grey junglefowl Gallus sonneratii Green junglefowl Gallus varius White-crested kalij pheasant Lophura l. hamiltoni Nepal Kalij pheasant Lophura l. leucomelana Crawfurd's kalij pheasant Lophura l. crawfurdi Lineated kalij pheasant Lophura l. lineata True silver pheasant Lophura n. nycthemera Berlioz’s silver pheasant Lophura n. berliozi Lewis’s silver pheasant Lophura n. lewisi Edwards's pheasant Lophura edwardsi edwardsi Vietnamese pheasant Lophura e. hatinhensis Swinhoe's pheasant Lophura swinhoii Salvadori's pheasant Lophura inornata Malaysian crestless fireback Lophura e. erythrophthalma Bornean crested fireback pheasant Lophura i. ignita/nobilis Malaysian crestless fireback/Vieillot's Pheasant Lophura i. rufa Siamese fireback pheasant Lophura diardi Southern Cavcasus Phasianus C. colchicus Manchurian Ring Neck Phasianus C. pallasi Northern Japanese Green Phasianus versicolor -

Complete Mitochondrial Genome of the Western Capercaillie Tetrao Urogallus (Phasianidae, Tetraoninae)
Zootaxa 4550 (4): 585–593 ISSN 1175-5326 (print edition) https://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2019 Magnolia Press ISSN 1175-5334 (online edition) https://doi.org/10.11646/zootaxa.4550.4.9 http://zoobank.org/urn:lsid:zoobank.org:pub:12E18262-0DCA-403A-B047-82CFE5E20373 Complete mitochondrial genome of the Western Capercaillie Tetrao urogallus (Phasianidae, Tetraoninae) GAËL ALEIX-MATA1,5, FRANCISCO J. RUIZ-RUANO2, JESÚS M. PÉREZ1, MATHIEU SARASA3 & ANTONIO SÁNCHEZ4 1Department of Animal and Plant Biology and Ecology, Jaén University, Campus Las Lagunillas, E-23071, Jaén, Spain. E-mail: [email protected] 2Departamento de Genética, Facultad de Ciencias, Universidad de Granada, Avda. Fuentenueva, 18071 Granada, Spain. 3BEOPS, 1 Esplanade Compans Caffarelli, 31000 Toulouse, France 4Department of Experimental Biology, Jaén University, Campus Las Lagunillas, E-23071, Jaén, Spain 5Corresponding author Gaël Aleix-Mata: [email protected] ORCID: 0000-0002-7429-4051 Francisco J. Ruíz-Ruano: [email protected] ORCID: 0000-0002-5391-301X Jesús M. Pérez: [email protected] ORCID: 0000-0001-9159-0365 Mathieu Sarasa: [email protected] ORDCID: 0000-0001-9067-7522 Antonio Sánchez: [email protected] ORCID: 0000-0002-6715-8158 Abstract The Western Capercaillie (Tetrao urogallus) is a galliform bird of boreal climax forests from Scandinavia to eastern Sibe- ria, with a fragmented population in southwestern Europe. We extracted the DNA of T. urogallus aquitanicus and obtained the complete mitochondrial genome (mitogenome) sequence by combining Illumina and Sanger sequencing sequence da- ta. The mitochondrial genome of T. urogallus is 16,683 bp long and is very similar to that of Lyrurus tetrix (16,677 bp). -

Western Capercaillie
WESTERN CAPERCAILLIE The grouse of the Today, it is an endangered species at Cansiglio namely due to the effects of the following factors: Cansiglio Forest • habitat loss, degradation and fragmentation due to lack of undergrowth, increase of natural A representative species of mountain and predators and climate changes; boreal ecosystems, the western capercaillie (Tetrao urogallus) is one of the species of grouse • diminishing undergrowth habitat resulting from inhabiting the Cansiglio Forest. deer grazing; • anthropic disturbances related to recreational Best known for the sensational displays of the activities. males during courtship, the western capercaillie lives in mature forests with large open areas and rich, yet not too thick, undergrowth, which Western capercaillie in the Cansiglio beechwood provides food, shelter and a place to nest in. (Photo: A. Ferro) In the Cansiglio Forest and other Rete Natura 2000 areas in Veneto, the periods in which tree cutting activities are allowed are compatible with this grouse’s breeding and rearing activities. Its presence is an assurance of quality for the environment, which is why improving its habitat means contributing to the increase of an area’s biodiversity. Western capercaillie (Photo: M. Bottazzo) Ideal habitat of the Western Capercaillie (Photo: R. Luise) Promuoviamo la Gestione Sostenibile delle Foreste PEFC/18-22-13/03 www.pefc.it Bio∆4 - “New instruments to enhance the biodiversity of cross-border forest ecosystems” ITAT2021 A project financed by the European Regional Development Fund (ERDF) within the context of the 2014 - 2020 Interreg V-A Italy - Austria programme (2017 call for tenders). The Italian portion is co-financed by the national rotating fund (C.I.P.E. -

G.A. TUNNICLIFFE, Canterbury Museum, Rolleston Avenue, Christchurch, New Zealand
MÄURI ORA, 1973, 1: 107-135 107 THE AVIFAUNA OF THE LAKE ELLESMERE AREA, CANTERBURY G.A. TUNNICLIFFE, Canterbury Museum, Rolleston Avenue, Christchurch, New Zealand. ABSTRACT The avifauna of the Lake Ellesmere area is described. A brief description of the area is given; past literature is reviewed; a checklist of birds, compiled from a variety of sources, is presented. For each species and subspecies (129 are listed), notes are given on Status and the generalised Vegetation zones or other habitat categories where it has been sighted or been found to breed. In one census area, lake level has a pronounced affect on which species are present. INTRODUCTION The avifauna of Lake Ellesmere has attracted little serious attention by ornithologists although its importance as a habitat for waterfowl and many other marsh dwelling species has been long recognized. Furthermore, there is a paucity of Information on the oceanic birds that occur on this part of the south Canterbury coastline. The early literature, e.g., Potts (1882), made only incid- ental reference to the birds of this area. Stead (1923), as ' Sibson in Falla et al (1966) previously indicated, first described the presence of several species of migrants previously unrecorded in New Zealand. From 1900 to 1929 Stead, at Ellesmere, made the first New Zealand sightings of the little whimbrel (Numenius minutus), Hudsonian godwit (Limosa haemastica), red-necked phalarope (Phalaropus lobatus), curlew sandpiper (Calidris ferruginea), red-necked Stint (Calidris ruficollis), pectoral sandpiper (Calidris melanotus) and sanderling (Calidris alba). Stead (1927, 1932) also investigated the Status, abundance, breeding activities and ecology of other species. -

Glucose Dehydrogenase Is Required for Normal Sperm Storage and Utilization in Female Drosophila Melanogaster Kaori Iida and Douglas R
The Journal of Experimental Biology 207, 675-681 675 Published by The Company of Biologists 2004 doi:10.1242/jeb.00816 Glucose dehydrogenase is required for normal sperm storage and utilization in female Drosophila melanogaster Kaori Iida and Douglas R. Cavener* Department of Biology, The Pennsylvania State University, 208 Mueller Laboratory, University Park, PA 16802, USA *Author for correspondence (e-mail: [email protected]) Accepted 28 November 2003 Summary Female sperm storage is a key factor for reproductive capsules of the mutant females. The storage defect was success in a variety of organisms, including Drosophila especially severe when the mutant females were crossed to melanogaster. The spermathecae, one of the Drosophila a Gld-mutant male that had previously mated a few hours sperm storage organs, has been suggested as a long-term before the experimental cross. Under this mating storage organ because its secreted substances may condition, the mutant females stored in the spermathecae enhance the quality of sperm storage. Glucose only one-third of the sperm amount of the wild-type dehydrogenase (GLD) is widely expressed and secreted in control females. In addition, the mutant females used the spermathecal ducts among species of the genus stored sperm at a slower rate over a longer period Drosophila. This highly conserved expression pattern compared with wild-type females. Thus, our results suggests that this enzyme might have an important role in indicate that GLD facilitates both sperm uptake and female fertility. Here, we examine the function of GLD in release through the spermathecal ducts. sperm storage and utilization using Gld-null mutant females. -

Slovenská Poľnohospodárska Univerzita V Nitre
SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV 1131583 BIOLOGICKÁ A EKOLOGICKÁ CHARAKTERISTIKA DRUHOV ČEĽADE PHASIANIDAE 2011 Jozef Čepišák SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV BIOLOGICKÁ A EKOLOGICKÁ CHARAKTERISTIKA DRUHOV PODČEĽADE PHASIANIDAE (Bakalárska práca) Študijný program: Udržateľné poľnohospodárstvo a rozvoj vidieka Študijný odbor: 4140700 Všeobecné poľnohospodárstvo Školiace pracovisko: Katedra environmentalistiky a zoológie Školiteľ: RNDr. Alena Rakovská, CSc. Nitra 2011 Jozef Čepišák ČESTNÉ VYHLÁSENIE Podpísaný, Jozef Čepišák vyhlasujem, že som záverečnú prácu na tému „Biologická a ekologická charakteristika druhov čeľade Phasianidae“ vypracoval samostatne s použitím uvedenej literatúry. Som si vedomý zákonných dôsledkov v prípade, ak hore uvedené údaje nie sú pravdivé. V Nitre 12.5.2011 ........................ podpis 3 Poďakovanie Touto cestou si dovoľujem poďakovať vedúcej bakalárskej práce RNDr. Alene Rakovskej, CSc. za jej pomoc, cenné rady a pripomienky pri spracovaní bakalárskej práce. 4 ABSTRAKT V predloženej bakalárskej práci „Biologická a ekologická charakteristika druhov čeľade Phasianidae“ je uvedená morfologická a anatomická stavba tela vtákov, spoločné znaky triedy vtáky (Aves) a ich fylogenetický pôvod. V práci uvádzame taxonómiu vybraných druhov bažantov, ktorá je podľa rôznych autorov rozdielna. Rad hrabavce (Galliformes) je bohatý na druhy, s charakteristickými vlastnosťami, ktoré sme bližšie špecifikovali. Stručne uvádzame i význam týchto vtákov pre ľudstvo a ich vzťah k človeku. Najväčšie druhové zastúpenie v rade hrabavcov má čeľaď bažantovité (Phasianidae). Do tejto čeľade patria hospodársky významne druhy hydiny, chované hlavne na mäsovú produkciu a vajcia. Patria sem aj naše pôvodné druhy vtákov (tetrov hlucháň, jarabica poľná, jariabok hôrny) a jeden nepôvodný druh, ktorým je bažant poľovný (Phasianus colchicus), súhrnne nazývané pernatá zver, ktorá je významná z poľovníckeho hľadiska. -

Full Page Photo
GEOLOGICA BALCANICA, 28. 1-2, Sofia, August. 1998, p. 53-60 Fossil birds of Dorkovo - an Early Pliocene site in the Rhodope Mts. (Southern Bulgaria) Zlatozar Boev National Museum of Natural History, Bulgarian Academy of Sciences, J()()(} Sofia, Bulgaria (Submitted 28.11.1997; accepted 12.02.1998) Iioes, 3. H. - HcKonaeMble nmUIIbl ~opKOBo U3 paHNefllluo Abstract. The avian fmds of the first in Bulgaria complexly lleHOBOi!o MeCmOHaxo:JK:OeHWI s PoOonax (IOxHDR Ii011zapW1). studied (1983-1985) palaeontological site of vertebrate fauna 0nHCb1BaiOTCII HaXO,llltH DTHU Hl nepsoro JtOMDneJtCHO are descnbed. Three fmds (distal endings of long bones of the Hccne.aosauuoro (1983-1985) a lionrapHH naneoaronorH<~ec legs) originated from the Early Pliocene (Ruscinian, MN xoro MeCTOH&XOli(JleHHJI DOJBOBO'IHOB $lyHhl. C paHBero DnH· zone 14, 5.4- 3.4 Ma). Taxonomically, they are referred to 3 oueua (pycuHHc"oro apyca, 5.4 - 3.4 MJIH. neT) o6aapy:&eHbl 3 families of Anseriformes and Galliformes: Balcanas plioca ,llHCT8nbHbiX cjlparMeHTa .llOnrHX ltOCTeii ltOHe'IHOCTeit 8 enica g. n. et sp. n. - humerus sin.; Tetrao rhodopensis sp. n. - TaJCCOHOMH'IeCJCOM OTHOWeHHH OHH OTHOCIITCll It 3 CCMeBCTB&M humerus sin. dist. and Perdicinae gen. indet. - tibiotarsus sin. OTplUlOB Anseriformes HGalliformes:Balcanas pliocaenica g.n. dist. Diagnosis ofBalcanas pliocaenica g. n. et sp. n.: A medium et sp. n. - humerus sin.; Tetrao rhodopensis sp. n. - humerus sin. sized anatid of Anatini tribe with narrower and more dist. H Perdicinae gen. indet. - tibiotarsus sin. dist. .l{HaraoJ transverally orientated fossa m. brachialis, obliquely outlined Balcanas pliocaenica g. n. et sp. n.: Cpe.aad no plllMepy aaa toward diaphysis medial edge of epicondylus ulnaris in ventral TH.ll HJ TPH6bl Anatini oTnH'I&IOillHHC.II: 6onee yJJtoii H none view, better developed fossa olecrani and shallower and pe'IHO OpHeHTHpOBaHHOB fossa m.