Ecosystem Effects of Mining Dr Matt Pinkerton
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CHECKLIST and BIOGEOGRAPHY of FISHES from GUADALUPE ISLAND, WESTERN MEXICO Héctor Reyes-Bonilla, Arturo Ayala-Bocos, Luis E
ReyeS-BONIllA eT Al: CheCklIST AND BIOgeOgRAphy Of fISheS fROm gUADAlUpe ISlAND CalCOfI Rep., Vol. 51, 2010 CHECKLIST AND BIOGEOGRAPHY OF FISHES FROM GUADALUPE ISLAND, WESTERN MEXICO Héctor REyES-BONILLA, Arturo AyALA-BOCOS, LUIS E. Calderon-AGUILERA SAúL GONzáLEz-Romero, ISRAEL SáNCHEz-ALCántara Centro de Investigación Científica y de Educación Superior de Ensenada AND MARIANA Walther MENDOzA Carretera Tijuana - Ensenada # 3918, zona Playitas, C.P. 22860 Universidad Autónoma de Baja California Sur Ensenada, B.C., México Departamento de Biología Marina Tel: +52 646 1750500, ext. 25257; Fax: +52 646 Apartado postal 19-B, CP 23080 [email protected] La Paz, B.C.S., México. Tel: (612) 123-8800, ext. 4160; Fax: (612) 123-8819 NADIA C. Olivares-BAñUELOS [email protected] Reserva de la Biosfera Isla Guadalupe Comisión Nacional de áreas Naturales Protegidas yULIANA R. BEDOLLA-GUzMáN AND Avenida del Puerto 375, local 30 Arturo RAMíREz-VALDEz Fraccionamiento Playas de Ensenada, C.P. 22880 Universidad Autónoma de Baja California Ensenada, B.C., México Facultad de Ciencias Marinas, Instituto de Investigaciones Oceanológicas Universidad Autónoma de Baja California, Carr. Tijuana-Ensenada km. 107, Apartado postal 453, C.P. 22890 Ensenada, B.C., México ABSTRACT recognized the biological and ecological significance of Guadalupe Island, off Baja California, México, is Guadalupe Island, and declared it a Biosphere Reserve an important fishing area which also harbors high (SEMARNAT 2005). marine biodiversity. Based on field data, literature Guadalupe Island is isolated, far away from the main- reviews, and scientific collection records, we pres- land and has limited logistic facilities to conduct scien- ent a comprehensive checklist of the local fish fauna, tific studies. -

The Importance of Krill Predation in the Southern Ocean Philip N Trathan
The importance of krill predation in the Southern Ocean Philip N Trathan and Simeon L Hill British Antarctic Survey, Madingley Road, Cambridge CB3 0ET 1 Abstract Antarctic krill is a major prey species for a diverse array of Southern Ocean predators. The amount of krill that predators consume, and how this changes over space and time, is a key issue in understanding both regional and circumpolar aspects of the Southern Ocean food- web. We assess current knowledge of consumption by the various predator groups, and the ecological processes through which krill and its predators influence each other. Knowledge has improved greatly over recent decades and has revealed a high level of complexity in the processes that govern krill consumption. We focus on the Antarctic Peninsula and Scotia Sea region where both krill and its consumers occur in significant concentrations and where an updated estimate of krill consumption by the main vertebrate groups is 55 million tonnes per year. Research has mainly focused on mammalian and avian predators of post-larval krill, particularly penguins. Potentially important consumer groups like fish, cephalopods and carnivorous zooplankton remain poorly understood, as does consumption of the early life stages of krill. As a consequence of these knowledge gaps and the variability that arises from complexity, a reliable seasonally, spatially or taxonomically resolved description of krill consumption remains elusive. One of the key motivations for attempting to estimate krill consumption is to understand how changes in krill availability impact predator populations. Such understanding is an important requirement for ecosystem based management of the Antarctic krill fishery. -

Updated Checklist of Marine Fishes (Chordata: Craniata) from Portugal and the Proposed Extension of the Portuguese Continental Shelf
European Journal of Taxonomy 73: 1-73 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.73 www.europeanjournaloftaxonomy.eu 2014 · Carneiro M. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:9A5F217D-8E7B-448A-9CAB-2CCC9CC6F857 Updated checklist of marine fishes (Chordata: Craniata) from Portugal and the proposed extension of the Portuguese continental shelf Miguel CARNEIRO1,5, Rogélia MARTINS2,6, Monica LANDI*,3,7 & Filipe O. COSTA4,8 1,2 DIV-RP (Modelling and Management Fishery Resources Division), Instituto Português do Mar e da Atmosfera, Av. Brasilia 1449-006 Lisboa, Portugal. E-mail: [email protected], [email protected] 3,4 CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected], [email protected] * corresponding author: [email protected] 5 urn:lsid:zoobank.org:author:90A98A50-327E-4648-9DCE-75709C7A2472 6 urn:lsid:zoobank.org:author:1EB6DE00-9E91-407C-B7C4-34F31F29FD88 7 urn:lsid:zoobank.org:author:6D3AC760-77F2-4CFA-B5C7-665CB07F4CEB 8 urn:lsid:zoobank.org:author:48E53CF3-71C8-403C-BECD-10B20B3C15B4 Abstract. The study of the Portuguese marine ichthyofauna has a long historical tradition, rooted back in the 18th Century. Here we present an annotated checklist of the marine fishes from Portuguese waters, including the area encompassed by the proposed extension of the Portuguese continental shelf and the Economic Exclusive Zone (EEZ). The list is based on historical literature records and taxon occurrence data obtained from natural history collections, together with new revisions and occurrences. -

120 South Georgian Diving-Petrel
Text and images extracted from Marchant, S. & Higgins, P.J. (co-ordinating editors) 1990. Handbook of Australian, New Zealand & Antarctic Birds. Volume 1, Ratites to ducks; Part A, Ratites to petrels. Melbourne, Oxford University Press. Pages 263-264, 719-724; plate 53. Reproduced with the permission of Bird life Australia and Jeff Davies. 263 Order PROCELLARIIFORMES A rather distinct group of some 80-100 species of pelagic seabirds, ranging in size from huge to tiny and in habits from aerial (feeding in flight) to aquatic (pursuit-diving for food), but otherwise with similar biology. About three-quarters of the species occur or have been recorded in our region. They are found throughout the oceans and most come ashore voluntarily only to breed. They are distinguished by their hooked bills, covered in horny plates with raised tubular nostrils (hence the name Tubinares). Their olfactory systems are unusually well developed (Bang 1966) and they have a distinctly musky odour, which suggest that they may locate one another and their breeding places by smell; they are attracted to biogenic oils at sea, also no doubt by smell. Probably they are most closely related to penguins and more remotely to other shorebirds and waterbirds such as Charadrii formes and Pelecaniiformes. Their diversity and abundance in the s. hemisphere suggest that the group originated there, though some important groups occurred in the northern hemisphere by middle Tertiary (Brodkorb 1963; Olson 1975). Structurally, the wings may be long in aerial species and shorter in divers of the genera Puffinus and Pel ecanoides, with 11 primaries, the outermost minute, and 10-40 secondaries in the Oceanitinae and great albatrosses respectively. -

Order MYCTOPHIFORMES NEOSCOPELIDAE Horizontal Rows
click for previous page 942 Bony Fishes Order MYCTOPHIFORMES NEOSCOPELIDAE Neoscopelids By K.E. Hartel, Harvard University, Massachusetts, USA and J.E. Craddock, Woods Hole Oceanographic Institution, Massachusetts, USA iagnostic characters: Small fishes, usually 15 to 30 cm as adults. Body elongate with no photophores D(Scopelengys) or with 3 rows of large photophores when viewed from below (Neoscopelus).Eyes variable, small to large. Mouth large, extending to or beyond vertical from posterior margin of eye; tongue with photophores around margin in Neoscopelus. Gill rakers 9 to 16. Dorsal fin single, its origin above or slightly in front of pelvic fin, well in front of anal fins; 11 to 13 soft rays. Dorsal adipose fin over end of anal fin. Anal-fin origin well behind dorsal-fin base, anal fin with 10 to 14 soft rays. Pectoral fins long, reaching to about anus, anal fin with 15 to 19 rays.Pelvic fins large, usually reaching to anus.Scales large, cycloid, and de- ciduous. Colour: reddish silvery in Neoscopelus; blackish in Scopelengys. dorsal adipose fin anal-fin origin well behind dorsal-fin base Habitat, biology, and fisheries: Large adults of Neoscopelus usually benthopelagic below 1 000 m, but subadults mostly in midwater between 500 and 1 000 m in tropical and subtropical areas. Scopelengys meso- to bathypelagic. No known fisheries. Remarks: Three genera and 5 species with Solivomer not known from the Atlantic. All Atlantic species probably circumglobal . Similar families in occurring in area Myctophidae: photophores arranged in groups not in straight horizontal rows (except Taaningichthys paurolychnus which lacks photophores). Anal-fin origin under posterior dorsal-fin anal-fin base. -

Differences in Backscattering Strength Determined at 120 and 38 Khz for Three Species of Antarctic Macroplankton
MARINE ECOLOGY PROGRESS SERIES Vol. 93: 17-24, 1993 Published February 23 Mar. Ecol. Prog. Ser. l Differences in backscattering strength determined at 120 and 38 kHz for three species of Antarctic macroplankton ' British Antarctic Survey, Natural Environment Research Council, High Cross, Madingley Road, Cambridge CB3 OET, United Kingdom Department of Zoology - University of Cambridge, Downing Street, Cambridge CB2 3EJ. United Kingdom ABSTRACT. The ability to acoust~callyseparate zooplankton specles is an important requirement for ecological studies and to improve b~omassestimates. In order to distlngulsh between Euphausia superba and other swarm-form~ngmacroplankters we used a dual frequency echo-sounder (120 and 38 kHz) and echo-integrator during a serles of Longhurst Hardy Plankton Recorder (LHPR)hauls near South Georg~aWe compared the acoustic parameter Mean Volume Backscattering Strength (MVBS) according to the equation: AMVBS (dB) = MVBS 120 kHz - MVBS 38 kHz. Mean values of AIMVBS for E. superba, Themlsto gaudichaudii and E. friglda were 4.6, 9.7 and 15.6 dB, respectively, and were significantly d~fferent,allowing the 3 species to be separdted acoustically. INTRODUCTION Conversely, high frequencies necessary to detect small species suffer from greater attenuation and conse- Acoustic methods have been used for fish stock esti- quently have a very short range. For fish studies, the mation and ecological studies for more than 20 yr (Sund most common frequencies vary from 12 to 200 kHz 1935, Greenlaw 1979, Johannesson & Mitson 1983).The (Clay & Medwin 1977, Farquhar 1977, Saville 1977, so-called echo-integration method has had a worldwide Genin et al. 1988, Eckmann 1991) while for studying application and the merit of using acoustics relative to zooplankton 20 kHz to more than 1 MHz have been nets has frequently been discussed (Greenlaw 1979, used (Clay & Medwin 1977, Pieper 1979, Sameoto Pieper & Holliday 1984, Everson & Bone 1986a). -

Biodiversity of Arctic Marine Fishes: Taxonomy and Zoogeography
Mar Biodiv DOI 10.1007/s12526-010-0070-z ARCTIC OCEAN DIVERSITY SYNTHESIS Biodiversity of arctic marine fishes: taxonomy and zoogeography Catherine W. Mecklenburg & Peter Rask Møller & Dirk Steinke Received: 3 June 2010 /Revised: 23 September 2010 /Accepted: 1 November 2010 # Senckenberg, Gesellschaft für Naturforschung and Springer 2010 Abstract Taxonomic and distributional information on each Six families in Cottoidei with 72 species and five in fish species found in arctic marine waters is reviewed, and a Zoarcoidei with 55 species account for more than half list of families and species with commentary on distributional (52.5%) the species. This study produced CO1 sequences for records is presented. The list incorporates results from 106 of the 242 species. Sequence variability in the barcode examination of museum collections of arctic marine fishes region permits discrimination of all species. The average dating back to the 1830s. It also incorporates results from sequence variation within species was 0.3% (range 0–3.5%), DNA barcoding, used to complement morphological charac- while the average genetic distance between congeners was ters in evaluating problematic taxa and to assist in identifica- 4.7% (range 3.7–13.3%). The CO1 sequences support tion of specimens collected in recent expeditions. Barcoding taxonomic separation of some species, such as Osmerus results are depicted in a neighbor-joining tree of 880 CO1 dentex and O. mordax and Liparis bathyarcticus and L. (cytochrome c oxidase 1 gene) sequences distributed among gibbus; and synonymy of others, like Myoxocephalus 165 species from the arctic region and adjacent waters, and verrucosus in M. scorpius and Gymnelus knipowitschi in discussed in the family reviews. -

A Review of Lanternfishes (Families: Myctophidae and Neoscopelidae)
Zoological Studies 40(2): 103-126 (2001) A Review of Lanternfishes (Families: Myctophidae and Neoscopelidae) and Their Distributions around Taiwan and the Tungsha Islands with Notes on Seventeen New Records John Ta-Ming Wang* and Che-Tsung Chen Graduate School of Fisheries Science, National Taiwan Ocean University, Keelung, Taiwan 202, R.O.C. Fax: 886-2-28106688. E-mail: [email protected] (Accepted December 20, 2000) John Ta-Ming Wang and Che-Tsung Chen (2001) A review of lanternfishes (Families: Myctophidae and Neoscopelidae) and their distributions around Taiwan and the Tungsha Islands with notes on seventeen new records. Zoological Studies 40(2): 103-126. Lanternfishes collected during 9 cruises from 1991 to 1997 were studied. The area sampled lies between 19°N and 25°N and 114°E and 123°E. The specimens collected in this area comprise 40 species belong to 16 genera, among which 17 species are first records. These first record species include Benthosema fibulatum, Bolinichthys supralateralis, Electrona risso, Hygophum proximum, H. reinhardtii, Lampadena anomala, Lobianchia gemellarii, Lampanyctus niger, L. turneri, L. tenuiformis, Myctophum asperum, M. aurolaternatum, M. nitidulum, M. spinosum, Notolychnus valdiviae, Notoscopelus caudispinosus, and N. resplendens. Among these, six species, Bolinichthys supralateralis, Electrona risso, Lampanyctus turneri, Lampadena anomala, Notolychnus valdiviae, and Notoscopelus caudispinosus, are first records for the South China Sea, and the species, Lampadena anomala is a new record for Asian oceans (Table 1). Four species (Triphoturus microchir, Diaphus diadematus, D. latus, and D. taaningi) were controversial in previous reports, so they are discussed in this study. Geographic distributions and localities of catches of all lanternfish species are shown on the maps (Figs. -

Amphipod-Based Food Web: Themisto Gaudichaudii Caught in Nets and by Seabirds in Kerguelen Waters, Southern Indian Ocean
MARINE ECOLOGY PROGRESS SERIES Vol. 223: 261–276, 2001 Published November 28 Mar Ecol Prog Ser Amphipod-based food web: Themisto gaudichaudii caught in nets and by seabirds in Kerguelen waters, southern Indian Ocean Pierrick Bocher1, 2, Yves Cherel1,*, Jean-Philippe Labat3, Patrick Mayzaud3, Suzanne Razouls4, Pierre Jouventin1, 5 1Centre d’Etudes Biologiques de Chizé, UPR-CNRS 1934, 79360 Villiers-en-Bois, France 2Laboratoire de Biologie et Environnement Marins, EA 1220 de l'Université de La Rochelle, 17026 La Rochelle Cedex, France 3Laboratoire d'Océanographie Biochimique et d’Ecologie, ESA 7076-CNRS/UPMC LOBEPM, Observatoire Océanologique, BP 28, 06230 Villefranche-sur-Mer, France 4Observatoire Océanologique, UMR-CNRS/UPMC 7621, Laboratoire Arago, 66650 Banyuls-sur-Mer, France 5Centre d’Ecologie Fonctionnelle et Evolutive, UPR-CNRS 9056, 1919 Route de Mende, 34293 Montpellier Cedex 5, France ABSTRACT: Comparing food samples from diving and surface-feeding seabirds breeding in the Golfe du Morbihan at Kerguelen Islands to concurrent net samples caught within the predator forag- ing range, we evaluated the functional importance of the hyperiid amphipod Themisto gaudichaudii in the subantarctic pelagic ecosystem during the summer months. T. gaudichaudii occurred in high densities (up to 61 individuals m-3) in the water column, being more abundant within islands in the western part of the gulf than at open gulf and shelf stations. The amphipod was a major prey of all seabird species investigated except the South Georgian diving petrel, accounting for 39, 80, 68, 59 and 46% of the total number of prey of blue petrels, thin-billed prions, Antarctic prions, common div- ing petrels and southern rockhopper penguins, respectively. -

Guide to the Coastal Marine Fishes of California
STATE OF CALIFORNIA THE RESOURCES AGENCY DEPARTMENT OF FISH AND GAME FISH BULLETIN 157 GUIDE TO THE COASTAL MARINE FISHES OF CALIFORNIA by DANIEL J. MILLER and ROBERT N. LEA Marine Resources Region 1972 ABSTRACT This is a comprehensive identification guide encompassing all shallow marine fishes within California waters. Geographic range limits, maximum size, depth range, a brief color description, and some meristic counts including, if available: fin ray counts, lateral line pores, lateral line scales, gill rakers, and vertebrae are given. Body proportions and shapes are used in the keys and a state- ment concerning the rarity or commonness in California is given for each species. In all, 554 species are described. Three of these have not been re- corded or confirmed as occurring in California waters but are included since they are apt to appear. The remainder have been recorded as occurring in an area between the Mexican and Oregon borders and offshore to at least 50 miles. Five of California species as yet have not been named or described, and ichthyologists studying these new forms have given information on identification to enable inclusion here. A dichotomous key to 144 families includes an outline figure of a repre- sentative for all but two families. Keys are presented for all larger families, and diagnostic features are pointed out on most of the figures. Illustrations are presented for all but eight species. Of the 554 species, 439 are found primarily in depths less than 400 ft., 48 are meso- or bathypelagic species, and 67 are deepwater bottom dwelling forms rarely taken in less than 400 ft. -

Themisto Amphipods in High-Latitude Marine Pelagic Food Webs
1 Predatory zooplankton on the move: 2 Themisto amphipods in high-latitude marine pelagic food webs 3 4 Charlotte Havermans*1, 2, Holger Auel1, Wilhelm Hagen1, Christoph Held2, Natalie Ensor3, Geraint Tarling3 5 1 Universität Bremen, BreMarE - Bremen Marine Ecology, Marine Zoology, 6 PO Box 330 440, 28334 Bremen, Germany 7 2 Alfred-Wegener-Institut Helmholtz-Zentrum für Polar- und Meeresforschung, 8 Am Handelshafen 12, 27568 Bremerhaven, Germany 9 3 Natural Environment Research Council, 10 High Cross Madingley Road, Cambridge, CB3 0ET, United Kingdom 11 12 *corresponding author 13 E-mail: [email protected] 14 Tel: +49 421 218 63037 15 ORCID ID: 0000-0002-1126-4074 16 https://doi.org/10.1016/bs.amb.2019.02.002 17 ABSTRACT 18 Hyperiid amphipods are predatory pelagic crustaceans that are particularly prevalent in high-latitude 19 oceans. Many species are likely to have co-evolved with soft-bodied zooplankton groups such as salps 20 and medusae, using them as substrate, for food, shelter or reproduction. Compared to other pelagic 21 groups, such as fish, euphausiids and soft-bodied zooplankton, hyperiid amphipods are poorly studied 22 especially in terms of their distribution and ecology. Hyperiids of the genus Themisto, comprising seven 23 distinct species, are key players in temperate and cold-water pelagic ecosystems where they reach 24 enormous levels of biomass. In these areas, they are important components of marine food webs, and 25 they are major prey for many commercially important fish and squid stocks. In northern parts of the 26 Southern Ocean, Themisto are so prevalent that they are considered to take on the role that Antarctic 1 27 krill play further south. -
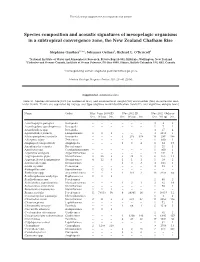
Marine Ecology Progress Series 503:23
The following supplement accompanies the article 1 Species composition and acoustic signatures of mesopelagic organisms in a subtropical convergence zone, the New Zealand Chatham Rise Stéphane Gauthier1, 2,*, Johannes Oeffner1, Richard L. O’Driscoll1 1National Institute of Water and Atmospheric Research, Private Bag 14-901, Kilbirnie, Wellington, New Zealand 2Fisheries and Oceans Canada, Institute of Ocean Sciences, PO Box 6000, Sidney, British Columbia V8L 4B2, Canada *Corresponding author: [email protected] Marine Ecology Progress Series: 503: 23–40 (2014) Supplement. Additional data Table S1. Species occurrence (Occ.) in number of tows, and abundance in weight (Wt) and number (No.) in successful mid - water trawls. Trawls are separated by voyage and type (daytime mark identification trawls ID, and nighttime oblique tows) Name Order May−June 2008 ID Nov 2011 ID Nov 2011 Oblique Occ. Wt (g) No. Occ. Wt (g) No. Occ. Wt (g) No. Acanthephyra pelagica Decapoda − − − − − − 3 9 3 Acanthephyra quadrispinosa Decapoda − − − − − − 1 7 1 Acanthephyra spp. Decapoda − − − − − − 3 17 4 Agrostichthys parkeri Lampriformes 1 8 1 − − − 3 1813 3 Alainopasiphaea australis Decapoda − − − 3 273 356 14 287 366 Allocyttus niger Zeiformes − − − − − − 1 600 1 Amphipod (unspecified) Amphipoda − − − 1 3 4 8 34 17 Anoplogaster cornuta Beryciformes − − − − − − 1 25 1 Apristurus spp. Carcharhiniformes − − − − − − 1 900 1 Argentina elongata Argentiniformes − − − − − − 4 541 6 Argyropelecus gigas Stomiiformes 2 20 2 2 38 5 8 165 12 Argyropelecus hemigymnus Stomiiformes 4 12 6 1 1 1 1 19 2 Astronesthes spp. Stomiiformes − − − 1 3 3 3 103 3 Atolla wyvillei Coronatae − − − − − − 4 83 7 Bathophilus ater Stomiiformes 1 12 1 − − − − − − Bathylagus spp.